Novocrania anomala and Protanthea simplex on sheltered circalittoral rock
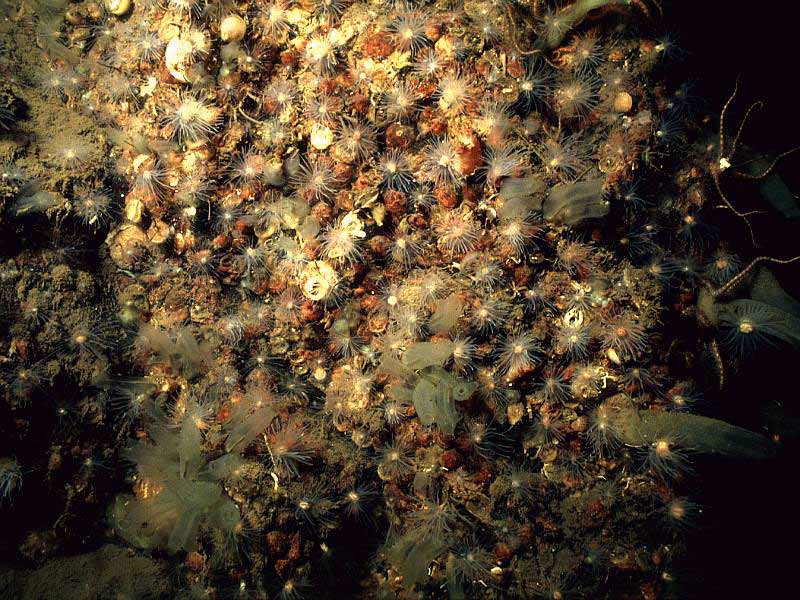
Circalittoral cliff face with dense brachiopods Neocrania anomala and Terebratulina retusa, the anemone Protanthea simplex and the ascidian Ciona intestinalis. Loch Duich, Highland.
Photographer: Sue Scott Copyright: Joint Nature Conservation Committee (JNCC)

Circalittoral cliff with Protanthea simplex, Sabella pavonina, Ciona intestinalis and Ophiothrix fragilis.
Photographer: Dr Keith Hiscock Copyright: Joint Nature Conservation Committee (JNCC)

Faunal dominated rock wall.
Photographer: Dr Keith Hiscock Copyright: Dr Keith Hiscock

| Researched by | John Readman, Angus Jackson, Kelsey Lloyd & Amy Watson | Refereed by | Admin |
|---|
Summary
UK and Ireland classification
Description
This biotope typically occurs in full to variable salinity conditions on very wave-sheltered circalittoral bedrock and boulder slopes subject to negligible tidal streams (this tends to be in the landward, very sheltered basins of fjordic sealochs). This biotope is characterized by often dense populations of the anemone Protanthea simplex, growing on the silty bedrock. The underlying rock surfaces are usually covered by encrusting red algae, the polychaete Spirobranchus triqueter, the brachiopods Novocrania anomala and Terebratulina retusa, the saddle oyster Pododesmus patelliformis and the polychaete Sabella pavonina. Scattered colonies of Alcyonium digitatum and the hydroid Bougainvillia ramosa may occasionally be recorded. A diverse range of ascidians including Ciona intestinalis, Ascidia mentula, Corella parallelogramma, Ascidia virginea, Polycarpa pomaria and Dendrodoa grossularia are also occasionally recorded. Echinoderms such as the common brittlestar Ophiothrix fragilis are frequently reported with their arms protruding from crevices in the rock, whilst the starfish Asterias rubens, Henricia oculata, and the sea urchin Echinus esculentus and Psammechinus miliaris are occasionally found on the boulder/rock surface. The whelk Buccinum undatum is often present but in very low numbers. The squat lobster Munida rugosa may be seen hiding in crevices. The hermit crab Pagurus bernhardus may also be recorded. (Information from Connor et al., 2004; JNCC, 2105).
Depth range
5-10 m, 10-20 m, 20-30 m, 30-50 mAdditional information
CR.LCR.BrAS.NeoPro includes two sub-biotopes (NeoPro.FS and NeoPro.VS) depending on the salintiy regime. Both are found in simialr physical habitats but Dendrodoa grossularia becomes common is the variable salinity sub-biotope (NeoPro.VS) and Protanthea simplex exhibits a higher abundance in the full salinty sub-biotope (NeoPro.FS) (JNCC, 2014).
Listed By
Habitat review
Ecology
Ecological and functional relationships
In this circalittoral biotope and similar sub-biotopes there are few algal species and limited primary production. The fauna is dominated by encrusting / attached species such as anemones, hydroids, brachiopods and solitary ascidians. The fauna consists predominantly of passive and active suspension feeders. There are a few errant predators and scavengers such as Cancer pagurus, and Asterias rubens. Particularly dense assemblages with abundant solitary sea squirts may provide shelter for other small fauna. Additional shelter may be provided by large sponges, particularly in the sub-biotopes.Seasonal and longer term change
Ciona intestinalis is typically an annual species, particularly in reduced salinity water although reproduction can occur throughout the year. There will be some temporal changes in the coverage and shelter provided by this species. Otherwise, it is unlikely that there will be any seasonal or other temporal changes in the biotope.Habitat structure and complexity
The sheltered conditions of the landward basins of sea lochs can allow quite species-rich assemblages to develop. Several tube worm species, including Chaetopterus variopedatus can be used by Protanthea simplex as a substratum. Larger species, including sponges and solitary ascidians provide greater spatial complexity. Higher densities of the hard, permanently attached Novocrania anomala may exclude other species.Productivity
Primary productivity in this biotope is low although coralline algae were recorded in more than half of the records of this biotope and the brown algae Pseudolithoderma extensum in approximately one quarter of all records (JNCC, 1999). However, quite high densities of encrusting or attached organisms, primarily brachiopods, sea loch anemones, solitary ascidians and tube worms can result in quite high secondary productivity through suspension feeding.Recruitment processes
Recruitment is primarily through pelagic larvae. Intense local recruitment can occur with Ciona intestinalis where sticky mucus strings containing eggs and larvae are trapped round nearby adults or other objects. Dispersal ability of Novocrania anomala may be limited and only occur from local populations. Protanthea simplex with is long-lived pelagic larval stage has considerable dispersal potential.Time for community to reach maturity
None of the main characterizing species are particularly long lived. Novocrania anomala survives for possibly up to 10 years. Ciona intestinalis lives typically only for a year and populations can reproduce throughout the year. Large sponges may have considerably greater longevity and slower growth. It may take over five years for the community to reach maturity.Additional information
-Preferences & Distribution
Habitat preferences
| Depth Range | 5-10 m, 10-20 m, 20-30 m, 30-50 m |
|---|---|
| Water clarity preferences | Poor clarity / Extreme turbidity, Very high clarity / Very low turbidity, See additional information |
| Limiting Nutrients | Data deficient |
| Salinity preferences | Full (30-40 psu) |
| Physiographic preferences | Enclosed coast or Embayment |
| Biological zone preferences | Circalittoral |
| Substratum/habitat preferences | Bedrock, Large to very large boulders, Small boulders |
| Tidal strength preferences | Very weak (negligible), Weak < 1 knot (<0.5 m/sec.) |
| Wave exposure preferences | Extremely sheltered, Sheltered, Very sheltered |
| Other preferences |
Additional Information
The two sub-biotopes included within this assessment are characterized by variable, reduced or low salinity which may influence biotope structure. The two sub-biotopes have some similarities although SCR.NeoPro.Den is more species rich and may occur in more open lochs (so far it has only been recorded from Loch Etive). The temperature preferences of the individual species selected to represent the biotope are quite different to the temperatures in which the biotope occurs in Britain and Ireland. For instance, Ciona intestinalis has a world-wide distribution and optimal growth occurs at between 15-20 degrees C, considerably higher than water temperatures on the west coast of Scotland. Protanthea simplex extends further north into colder waters. No information is available regarding limiting nutrients.Species composition
Species found especially in this biotope
Rare or scarce species associated with this biotope
-
Additional information
The biotope assessment also covers two sub-biotopes. Ciona intestinalis is not one of the characterizing species in SCR.NeoPro.CaTw. However, Ciona intestinalis is assumed to form a suitable surrogate, representing the sensitivity of the various other solitary ascidians in the sub-biotope.Sensitivity review
Sensitivity characteristics of the habitat and relevant characteristic species
CR.LCR.BrAs.NovPro and its sub-biotopes CR.LCR.BrAs.NovPro.FS and CR.LCR.BrAs.NovPro.VS occur on steep or vertical sheltered bedrock and boulder slopes in the circalittoral, typically in fjordic sealochs. CR.LCR.BrAs.NovPro experiences full or variable salinity, and its two sub-biotopes differ in salinity; the low to full salinity NovPro.VS (salinity between <18-35 psu) and the full salinity NovPro.FS (salinity of between 30-35 psu).
The brachiopod Novocrania anomala (previously Neocrania anomala) is the characterizing species for this group. The anemone Protanthea simplex is a characterizing species of the full salinity NovPro.FS, but is only occasionally seen in the variable salinity NovPro.VS. In the variable salinity sub-biotope (NovPro.VS), a diverse range of ascidians, including Dendrodoa grossularia, Ciona intestinalis and Ascidia mentula become more dominant. Sarcodictyon roseum is present in the CR.LCR.BrAs.NovPro.VS, although the description and species list only record species as occasionally present and is considered where appropriate. Other species present in these biotopes are considered transient, mobile or ubiquitous and are therefore not considered significant to assessment of the sensitivity of these biotopes. Sensitivity assessments therefore focus on Novocrania anomala, Protanthea simplex and the ascidians (including Dendrodoa grossularia, Ciona intestinalis and Ascidia mentula).
Resilience and recovery rates of habitat
Novocrania anomala (previously Neocrania anomala) is an inarticulate brachiopod that cements its lower shell to the hard substrata (and is therefore sessile) following a pelagic larval stage (Nielsen, 1991, Alvarez & Emig 2000). Brachiopods tend to grow quickly initially, in order to increase survival in the early, most vulnerable life stage. Growth subsequently becomes more stable, diminishing in the latest stages. In general, brachiopods to live up to about 10 years (Alvarez & Emig, 2000). Novocrania anomala possesses no pedicle, cementing its ventral valve directly to the substratum instead and orientating with the dorsal side up, the anterior margin directed upwards away from the substratum (Ruppert & Barnes, 1994). The species is free-spawning and fertilization is external in the surrounding water column. The eggs are denser than seawater and hatch into a free-swimming larval stage. The larvae are fully developed within three days and settle out in no more than a few days, limiting the dispersal range. Although the species may inhabit areas with water flow rates of up to 3 knots, it is often restricted to sheltered habitats such as sea lochs, which may reduce dispersal ability (Jackson, 2000). No information was available about fecundity. Reproduction occurs annually and over an extended period of time (Long & Stricker, 1991; James et al., 1992) with spawning reported between April and October in the South of France and Scotland (Joubin, 1886; Rowell, 1960 cited in James et al., 1992). Novocrania anomala is also capable of recovery from considerable damage to the shell and soft tissue, the adults can be maintained quite well in aquaria and are generally hardy organisms (James et al., 1992).
Protanthea simplex is a small (2 cm high), delicate anemone (Jackson, 2008) commonly found in Scottish lochs (Wood, 2005), which appears to be the southern limit to this species’ distribution (NBN, 2016), although one record exists off Connemara, Ireland (Seeley, 2006). This anemone is sociable and beds of up to 2000 per m2 are found in Scandinavia (Wood, 2005). Protanthea simplex has a long-lived pelagic larval stage (15-20 days, at 10-12°C, in the plankton before settling) and therefore has considerable dispersal potential, with breeding taking place between September and October in Sweden (Jackson, 2008). Fragments of tissue in this species (except the tentacles) are capable of regenerating into complete anemones, a form of vegetative, asexual reproduction (Manuel, 1988). Anemones are not completely sessile and are capable of slow movement. For example, Sebens (1981) observed immigration to cleared patches of the long-lived anemone Anthopleura xanthogrammica as the primary driver towards recovery. Sebens (1981) cleared intertidal patches of Anthopleura xanthogrammica at Mukkaw Bay, WA observing the effects over four years. Even after four years, cleared areas were not back to pre-removal population densities. Chia & Spaulding (1972) studying the anemone Tealia crassicornis found no sign of gonad development at 14 months old.
Solitary ascidians are discrete creatures which do not fuse with others (unlike colonial ascidians), but may still form dense beds (e.g. up to 5000 individuals/m² for Ciona intestinalis) (Naylor, 2011). Dendrodoa grossularia, Ascidia mentula and Ciona intestinalis occur across the western, northern and southern coasts of the UK, with more scattered records on the eastern coast (NBN, 2015). Dendrodoa grossularia is a small solitary ascidian, 1.5-2 cm diameter (Millar, 1954). Settlement occurs from April-June, individuals reach their maximum size by the following summer. Life expectancy is expected to be 18-24 months. Sexual maturity is reached about one year after larval settlement and the release of gametes occurs from spring-autumn, with peaks in early spring and another in late summer. Gamete release is reduced at temperatures above 15°C and totally suppressed above ca 20°C (Millar, 1954). Kenny & Rees (1994) observed Dendrodoa grossularia was able to recolonize rapidly following aggregate dredging. Following experimental dredging of a site off the English coast, which extracted an area of 1-2 m wide and 0.3-0.5 m deep, Dendrodoa grossularia was able to recolonize and attained 40% of pre-dredge abundance and 23% of biomass within eight months. This recovery rate combined with the ability of this species to reach sexual maturity within its first year suggests that Dendrodoa grossularia can recover from disturbance events within 2 years. The species has a life expectancy of 18-24 months so that abundant populations are maintained by regular annual recruitment.
In Ciona intestinalis, spawning has been reported as year-round in temperate conditions (MBA, 1957, Yamaguchi, 1975, Caputi et al., 2015) with seasonal spawning observed in colder climates from May to June on the Canadian coast (Carver et al., 2006) and in shallower habitats in Sweden (Svane & Havenhand, 1993). Oviparous solitary ascidians generally spawn both oocytes and sperm into the water column, where the resultant fertilized eggs develop into free-swimming, non-feeding larvae. The eggs are negatively buoyant and slightly adhesive and are either released freely or in mucus strings that are especially adhesive. These strings have a tendency to settle close to or on the parent ascidian. In vitro studies conclude that fertilization proceeds normally whether in the water column or attached to the mucus string. The hatched free-swimming larvae settle nearby, are held by the mucus string until settlement or escape as plankton. Retention in the mucus string may explain the dense aggregations of adults found (Svane & Havenhand, 1993). In vitro studies indicate that both spawning and settlement are controlled by light. However, Ciona intestinalis has been observed in vivo to spawn and settle at any time of the day (Whittington, 1967; Svane & Havenhand, 1993). In the Mediterranean, population collapses of Ciona intestinalis were observed, followed by recovery within 1-2 years (Caputi et al., 2015). The collapses are still poorly understood, although low salinity (Pérès, 1943) and temperature (Sabbadin, 1957) were suggested as possible drivers.
Ascidia mentula is a larger (up to 18 cm long) and longer lived (up to seven years) ascidian compared to Ciona intestinalis (Rowley, 2008). Recruitment was reported to occur year round in Sweden at depths greater than 20 m, with seasonal spawning occurring at 15 m (where sea temperature variability is much greater). Both active larvae settlement distribution and passive deposition of larvae (i.e. purely hydrodynamic processes) have been proposed (Havenhand & Svane, 1991 see also Meadows & Campbell, 1972; Scheltema, 1974; Butman, 1987). Long-term data from populations of the ascidian Ascidia mentula on subtidal vertical rock indicated that recruitment of Ascidia mentula larvae was positively correlated with adult population density, and then by subsequent active larval choice at smaller scales. Factors influencing larval settlement have been listed as light, substratum inclination and texture (Havenhand & Svane, 1989). Sebens (1985, 1986) described the recolonization of epifauna on vertical rock walls. Rapid colonizers such as encrusting corallines, encrusting bryozoans, amphipods and tubeworms recolonized within 1-4 months. Ascidians such as Dendrodoa carnea, Molgula manhattensis and Aplidium spp. achieved significant cover in less than a year, and, together with Halichondria panicea, reached pre-clearance levels of cover after two years. A few individuals of Alcyonium digitatum and Metridium senile colonized within four years (Sebens, 1986) and would probably take longer to reach pre-clearance levels.
Sarcodictyon roseum forms small colonies on rock surfaces and occasionally on shell, with polyps that reach up to 1 cm in height. It is found from the intertidal to ca 100m and from the Mediterranean to the North Sea (Bay-Nouailhat, 2007; Hayward & Ryland, 1995b). It is found across the western coasts of the British Isles from the Channel Isles to the north coast of Scotland (NBN, 2016) and, more widely, from the Mediterranean to Sweden (Dyntaxa, 2013; van Ofwegen et al., 2001, cited from van Ofwegen, 2015). Very little information is available for this species.
Resilience assessment. The ascidians are likely to recover from any level of mortality relatively quickly, however, Novocrania anomala and Protanthea simplex are longer lived, slower growing and are unlikely to fully recover from significant mortality within two years. When resistance is None or Low, resilience is classed as ‘Medium’, but when resistance is Medium or High, resilience is classed as ‘High’.
Hydrological Pressures
Use [show more] / [show less] to open/close text displayed
| Resistance | Resilience | Sensitivity | |
Temperature increase (local) [Show more]Temperature increase (local)Benchmark. A 5°C increase in temperature for one month, or 2°C for one year. Further detail EvidenceNovocrania anomala extends as far south as the Mediterranean. Radolović et al. (2015) describe the gregarious settlement of Novocrania anomala in a cave in the Adriatic coast of Croatia. Despite this geographical range, Hiscock (1985 cited in Hartnoll, 1998), noted that seasonal shallow thermoclines may form, particularly in sheltered areas such as sea lochs, and extend down to 15 m. Some animals such as the brachiopods Crania (syn. Novocrania) and Terebratulina seem restricted to below this thermocline. It is, therefore, possible that geographical populations may acclimate to local temperature (Hartnoll, 1998). Protanthea simplex is a northerly species, with its southern distribution limit typically in Scotland, although one record exists off Connemara, Ireland (Seeley, 2006). Protanthea simplex experienced a marked long-term decline in Swedish and Norwegian sites between 1972 and 1982, experiencing ca 50% mortality (density of ca 600 to ca 300) at 20 m and virtual population collapse at 15 m (reduction in density from 400-500 to <50) over ten years (Heip et al., 1985). This decline appeared to coincide with a warm period between 1973 and 1978 of up to ca +2°C deviation and an increase in Novocrania anomala (Heip et al., 1985). Ciona intestinalis is considered a cold water or temperate species but has been found as far south as Cape Verde, although these tropical populations are likely transitory (Monniot & Monniot, 1994). Temperature tolerance varies among geographical populations or ecotypes. Adult Ciona intestinalis is reported as tolerant of temperatures up to 30°C (Dybern, 1965; Therriault & Herborg, 2008), although Petersen & Riisgard (1992) noted that filtration rates declined above 21°C, which suggested thermal stress, and indicated that long-term survival was likely to require temperatures lower than the 30°C. Other studies also indicated that Ciona intestinalis exhibits a decline in ammonia excretion rate and oxygen consumption rate above 18°C (Zhang and Fang 1999, Zhang et al., 1999). The effect of higher temperatures on Ascidia mentula is not as well researched. It is distributed from Norway through to the Mediterranean and the Black Sea, and the species appears to tolerate a broad range of temperatures. Svane (1984) found that in Sweden, whilst lower temperatures decreased recruitment, populations responded positively to the “warm period” of 1972-1976 (Glantz, 2005), with an increase in population density across all sites in the study and a gradual decrease during the ensuing “cold period”, and minor fluctuations throughout. Unusually high mean temperatures in 1975 did result in higher recruitment, with colder temperatures in January 1976 and spring 1979 coinciding with very little recruitment. Svane (1984) found that, unlike recruitment, mortality was regulated locally and independent of temperature within the range of the study (mean monthly deviation of ±3°C)(Svane, 1984). Sensitivity assessment. Whilst Novocrania anomala and the ascidians are unlikely to be affected by a temperature increase at the benchmark level, Protanthea simplex, already at its southern-most limit, is unlikely to tolerate a long-term increase in temperature e.g. 2°C for a year. Population collapse has been recorded at 15 m in Sweden during an extended warm period (longer than the benchmark level). Resistance has therefore been assessed as ‘Low’, Resilience has been assessed as ‘Medium’ and sensitivity is therefore recorded as ‘Medium’. | LowHelp | MediumHelp | MediumHelp |
Temperature decrease (local) [Show more]Temperature decrease (local)Benchmark. A 5°C decrease in temperature for one month, or 2°C for one year. Further detail EvidenceThe characterizing brachiopod Novocrania anomala has been recorded as far north as Svalbard (Greig, 1924, cited in Prestrud et al., 2004) and the anemone Protanthea simplex is a northerly species, occurring in Scandinavia and recorded in across Scotland (Wood, 2005; NBN, 2015). A reduction in temperature would probably be beneficial and could result in distribution expansion of Protanthea simplex. Tolerance for low temperatures varies among geographical populations of ascidians. In the Mediterranean, most adult Ciona intestinalis die when temperatures fall below 10°C, and the population is maintained by the survival of younger individuals, which are more tolerant of colder temperatures (Marin et al., 1987). Observation of Scandinavian populations indicated a higher mortality rate of Ciona intestinalis during the coldest period of the year (temperatures down to 1°C) (Dybern, 1965). In Scandinavian populations, normal egg development requires 8-22°C and larval development occurs between 6-24°C (Dybern, 1965). Larval temperature tolerances may play a part in successful recruitment in unseasonable temperature fluctuations. Ciona savigny larvae were found to acclimate to temperature, with embryos collected in the summer dividing normally between 14 - 27°C and embryos collected in the winter dividing normally between 10 - 20°C (Nomaguchi et al., 1997). Ascidia mentula is distributed from Norway through to the Mediterranean and the Black Sea, and the species appears to tolerate a broad range of temperatures. Svane (1984) found that in Sweden, whilst lower temperatures (of ±3°C of the monthly mean) decreased recruitment, mortality did not significantly increase. Shallow populations (15 m) experiencing much greater seasonal variability did exhibit seasonal spawning rather than year-round spawning that occurs in more temperate and deeper populations (Svane, 1984). Populations responded positively to the ‘warm period’ of 1972-1976 (Glantz, 2005), with an increase in population density across all sites in the study and a gradual decrease during the ensuing ‘cold period’, with minor fluctuations throughout. Unusually high mean temperatures in 1975 did result in higher recruitment, with colder temperatures in January 1976 and spring 1979 coinciding with very little recruitment. Svane (1984) found that, unlike recruitment, mortality was regulated locally and independent of temperature within the range of the study (mean monthly deviation of ±3°C). Sensitivity assessment. None of the characterizing species for this biotope are at their northern distribution limit and are unlikely to be affected by a reduction in temperature at the benchmark level. Resistance is therefore assessed as ‘High’, Resilience as ‘High’ and Sensitivity is therefore recorded as ‘Not Sensitive’. | HighHelp | HighHelp | Not sensitiveHelp |
Salinity increase (local) [Show more]Salinity increase (local)Benchmark. A increase in one MNCR salinity category above the usual range of the biotope or habitat. Further detail EvidenceNovocrania anomala occurs in biotopes ranging across all salinity regimes – from Low (<18 ppt) through to Full (30-35 ppt). No literature could be found relating to this brachiopod in hypersaline conditions. Protanthea simplex prefers full salinity habitats over lower salinity as, whilst it is characterizing in NovPro.FS, it is only occasionally found in the variable salinity biotope NovPro.VS (Connor et al., 2004). No evidence could be found for the effect of hypersaline conditions. Ciona intestinalis has been classified as euryhaline with a high salinity tolerance range (12-40‰) although it typically occurs in full salinity conditions (>30‰) (Tillin & Tyler-Walters, 2014). Ciona intestinalis has been found in salinities ranging from 11 to 33 PSU in Sweden, although the same study found that parent acclimation to salinity (high or low) has an overriding and significant effect on larval metamorphic success, independent of parent origins (Renborg, 2014). Sensitivity assessment. NovPro and NovPro.FS are found in full salinity, but 'No evidence' for the characterizing species in hypersaline conditions could be found. | No evidence (NEv)Help | Not relevant (NR)Help | No evidence (NEv)Help |
Salinity decrease (local) [Show more]Salinity decrease (local)Benchmark. A decrease in one MNCR salinity category above the usual range of the biotope or habitat. Further detail EvidenceNovocrania anomala occurs in biotopes ranging across all salinity variants, from Low (<18 ppt) through to full (30-35 ppt). Whilst Novocrania anomala and the ascidians are likely to be unaffected by a decrease in salinity at the benchmark level, the reduced presence of Protanthea simplex from the variable salinity NovPro.VS indicates that it is at least partially intolerant of salinity decrease. It should also be noted that, given the biotope descriptions, a change from NovPro.FS to NovPro.VS would occur in the event of a permanent salinity reduction at the benchmark level. Whilst no evidence could be found directly for Novocrania anomala, Hammond (1983) reported that Lingula anatina could tolerate salinities ranging from 20‰ to 50‰ for prolonged periods (more than four weeks) and survived in salinities as low as 5‰ for short periods of time (snap response failed at 1 day) and 11.5‰ for 19 days. Other studies report tolerances of ca 16 to 18‰ (Emig, 1997 and references therein) reported more conservative tolerance. It should be noted that the ability for brachiopods to respire anaerobically within their closed shell would enable them to survive short-term changes (James et al., 1992). Ciona intestinalis has been classified as euryhaline with a high salinity tolerance range (12-40‰) although it typically occurs in full salinity conditions (>30‰) (Tillin & Tyler-Walters, 2014) but has been found in Scandinavian waters in salinities as low as 11 PSU (Renborg, 2014, Dybern, 1967). Adult acclimation to salinity was shown to have an overriding and significant effect on larval metamorphic success, independent of parent origins (Renborg, 2014). ‘Massive die-offs’ of Ciona intestinalis were observed following winter rains in Californian harbours with ‘massive recolonizations usually following in the spring’ (Lambert & Lambert, 1998). Population collapses of Ciona intestinalis in the Mediterranean have also been reported, and whilst the drivers for these events are not well understood, it has been postulated that low salinity could play a part (Péres, 1943; Caputi et al., 2015). Oxygen consumption rate has been shown to decline with decreasing salinity and ceased at 19‰ with siphons tightly closed. (Shumway, 1978). Ascidia mentula is found on the West coast of Norway in salinities greater than 20‰ (Dybern, 1969) and found in a brackish lake in Corsica with a salinity gradient of 6.5 to 18.5‰ Cl- (Verhoeven, 1978). Dendrodoa was observed as one of the dominant species in a study area in Lübeck Bay, Norway, where salinity was recorded as between 11.1 -15.0‰ (Gulliksen, 1977). Sensitivity assessment. The abundance of Protanthea simplex is reduced from NovPro.FS (Full salinity of 35 ppt) to NovPro.VS (Low- Variable salinity, <18– 35ppt) while the abundance of Dendrodoa grossularia increases (JNCC, 2014). Therefore, resistance is assessed as ‘Medium, resilience as ‘High’ and sensitivity as ‘Low’. | MediumHelp | HighHelp | LowHelp |
Water flow (tidal current) changes (local) [Show more]Water flow (tidal current) changes (local)Benchmark. A change in peak mean spring bed flow velocity of between 0.1 m/s to 0.2 m/s for more than one year. Further detail EvidenceThe CR.LCR.BrAs.NovPro biotope exists in weak to negligible tidal flow conditions (<0.5 m/sec.) Connor et al. (2004). High flow rates may reduce the abundance of Novocrania anomala and Protanthea simplex, although Novocrania anomala may inhabit areas with water flow rates of up to 3 knots (Jackson, 2000; 2008b). Decreases in water flow are unlikely to have any effect but increases in flow rate above weak may prevent the animals from maintaining posture and interfere with feeding. Increased flow rates may also sweep individuals off the substratum. As sessile filter feeders, ascidians generally require a reasonable water flow rate in order to ensure sufficient food availability. It was shown that in stagnant water, phytoplankton density became reduced in a 20-30 cm layer immediately above a dense colony of Ciona intestinalis (Riisgård et al., 1996). However, Ciona intestinalis has been recognised as tolerant of low water flow environments which it uses as a competitive advantage in areas with minimal water exchange and renewal such as harbours, marinas and docks, (Carver et al., 2006). Whilst Ciona intestinalis is typically found in areas of low flow, it can reportedly withstand flow rates up to 3 knots (1.5 m/s) (Jackson 2008b). If dislodged, juveniles and adults have a limited capability to re-attach, given calm conditions and prolonged contact with the new substrata (Millar 1971; Carver et al., 2006; Jackson, 2008). Hiscock (1983) found that, for the solitary ascidian Ascidia mentula, siphons closed when current velocity rose above about 0.15 m/s. Sensitivity assessment. Significant increases in water flow are likely to negatively affect the characterizing species, but mortality at the benchmark level is unlikely. Therfore, resistance is assessed as ‘High’, resilience as ‘High’ and the biotope is considered ‘Not sensitive’ at the pressure benchmark. | HighHelp | HighHelp | Not sensitiveHelp |
Emergence regime changes [Show more]Emergence regime changesBenchmark. 1) A change in the time covered or not covered by the sea for a period of ≥1 year or 2) an increase in relative sea level or decrease in high water level for ≥1 year. Further detail EvidenceChanges in emergence are not relevant to this biotope as it is restricted to fully subtidal/circalittoral conditions - the pressure benchmark is relevant only to littoral and shallow sublittoral fringe biotopes. | Not relevant (NR)Help | Not relevant (NR)Help | Not relevant (NR)Help |
Wave exposure changes (local) [Show more]Wave exposure changes (local)Benchmark. A change in near shore significant wave height of >3% but <5% for more than one year. Further detail EvidenceThe NovPro biotope complex is found in sheltered to extremely sheltered wave exposure conditions. Novocrania anomala is unlikely to tolerate wave action and is found in sheltered. Increases in wave exposure above moderately exposed would probably cause death (Jackson, 2000). High energy wave action can be detrimental to ascidian populations. This is mainly through physical damage to the sea squirts and through the abrasive action of suspended sediment (Jackson, 2008). Ciona intestinalis is often dominant in highly sheltered areas such as harbours (Carver et al., 2006). Decreases in wave exposure are unlikely to have any effect. If dislodged, juvenile and adult Ciona intestinalis have a limited capability to re-attach, given calm conditions and prolonged contact with the new substratum (Millar 1971; Carver et al., 2006; Jackson 2008;) but increases in wave exposure above moderately exposed are likely to cause a proportion of the population to die, especially in the shallower examples of the biotope if the cobbles and pebbles on which the biotope occurs are mobilized by wave action. Ascidia mentula has rarely been recorded at depths shallower than 15 m (Svane, 1984), it is possible that damage could occur if subjected to increased wave exposure. Sensitivity assessment. Whilst the characterizing species are likely to be affected by an increase in wave exposure, the NovPro biotope complex is found in sheltered to extremely sheltered wave exposure conditions and a change at the benchmark level is unlikely to increase mortality. Resistance is therefore assessed as ‘High’, with resilience as ‘High’ and the biotope is therefore assessed as ‘Not sensitive’ at the benchmark level. | HighHelp | HighHelp | Not sensitiveHelp |
Chemical Pressures
Use [show more] / [show less] to open/close text displayed
| Resistance | Resilience | Sensitivity | |
Transition elements & organo-metal contamination [Show more]Transition elements & organo-metal contaminationBenchmark. Exposure of marine species or habitat to one or more relevant contaminants via uncontrolled releases or incidental spills. Further detail EvidenceMercier et al. (1998) studied response to TBT exposure of temperate anemones (Metridium senile and Bunodactis stella), which metabolised and regulated butyl-tin uptake, and accumulated less than mussels (Fent, 1996) or symbiotic anemones (Mercier et al., 1996). Regulation of butyl-tin compounds in temperate sea anemones seemed to parallel that of trace metals (Mercier et al., 1998). Nevertheless, this pressure is Not assessed. | Not Assessed (NA)Help | Not assessed (NA)Help | Not assessed (NA)Help |
Hydrocarbon & PAH contamination [Show more]Hydrocarbon & PAH contaminationBenchmark. Exposure of marine species or habitat to one or more relevant contaminants via uncontrolled releases or incidental spills. Further detail EvidenceThis pressure is Not assessed but evidence is presented where available. CR.LCR.BrAs.NovPro is a sub-tidal biotope (Connor et al., 2004). Oil pollution is mainly a surface phenomenon its impact upon circalittoral turf communities is likely to be limited. However, as in the case of the Prestige oil spill off the coast of France, high swell and winds can cause oil pollutants to mix with the seawater and potentially negatively affect sublittoral habitats (Castège et al., 2014). Filter feeders are highly sensitive to oil pollution, particularly those inhabiting the tidal zones that experience high exposure and show correspondingly high mortality, as are bottom dwelling organisms in areas where oil components are deposited by sedimentation (Zahn et al., 1981). Smith (1968) studied the aftermath of the Torrey Canyon oil spill and subsequent use of dispersants in that area. The beadlet anemone Actinia equine and the dahlia anemone Tealia feline were considered some of the most resistant animals on the shore, being commonly found alive, and on 26th April they were found in pools between the tide-marks which appeared to be devoid of all other animals. Some Anemonia sulcata, Cylista elegans and Cereus pedunculatus were found dead; few survived. Little evidence for brachiopod resistance to hydrocarbon contamination could be found. | Not Assessed (NA)Help | Not assessed (NA)Help | Not assessed (NA)Help |
Synthetic compound contamination [Show more]Synthetic compound contaminationBenchmark. Exposure of marine species or habitat to one or more relevant contaminants via uncontrolled releases or incidental spills. Further detail EvidenceThis pressure is Not assessed but evidence is presented where available. | Not Assessed (NA)Help | Not assessed (NA)Help | Not assessed (NA)Help |
Radionuclide contamination [Show more]Radionuclide contaminationBenchmark. An increase in 10µGy/h above background levels. Further detail EvidenceNo evidence | No evidence (NEv)Help | Not relevant (NR)Help | No evidence (NEv)Help |
Introduction of other substances [Show more]Introduction of other substancesBenchmark. Exposure of marine species or habitat to one or more relevant contaminants via uncontrolled releases or incidental spills. Further detail EvidenceThis pressure is Not assessed. | Not Assessed (NA)Help | Not assessed (NA)Help | Not assessed (NA)Help |
De-oxygenation [Show more]De-oxygenationBenchmark. Exposure to dissolved oxygen concentration of less than or equal to 2 mg/l for one week (a change from WFD poor status to bad status). Further detail EvidenceThe brachiopod Terebratulina unguicula is dominant in a fjord that shows annual upwelling of anoxic waters; it is one of the few animals found in areas with oxygen levels frequently below 0.1 ml/l (Tunnicliffe & Wilson, 1988). The inarticulate Crania californica has a restricted distribution that may be a function of low dispersal. Over a number of years, one monitored brachiopod population was stable although a gradual retreat from the low oxygen waters was observed (Tunnicliffe & Wilson, 1988). The brachiopod Terebratulina septentionalis has been shown to survive in anoxic conditions for 3.5 days (through anaerobic respiration), however, its activity may be reduced (Hammen, 1977, cited in James et al., 1992). Thayer (1981) found that the brachiopods L. californianus and Terebratulina unguicula survived more than two weeks in anoxic conditions. Ellington(1982) reviewed the effects of hypoxia and anoxia on anemones and noted that survival was several days to weeks depending on species. Mazouni et al. (2001) noted that whilst oysters (Magallana gigas) can survive short-term exposure to periods of anoxia (Thau Lagoon, France), the associated biofouling community dominated by Ciona intestinalis suffered heavy mortality. It should be noted, however, that this species is frequently found in areas with restricted water renewal where oxygen concentrations may drop (Carver et al., 2006). While adverse conditions could affect health, feeding, reproductive capability and could eventually lead to mortality, recovery should be rapid. In general, respiration in most marine invertebrates does not appear to be significantly affected until extremely low concentrations are reached. For many benthic invertebrates, this concentration is about 2 ml/l (Herreid, 1980; Rosenberg et al., 1991; Diaz & Rosenberg, 1995). Cole et al. (1999) suggest possible adverse effects on marine species below 4 mg/l and probable adverse effects below 2 mg/l. The ability of solitary ascidians to withstand decreasing oxygen levels has not been well documented. Hiscock & Hoare (1975) reported an oxycline forming in the summer months (Jun-Sep) in a quarry lake (Abereiddy, Pembrokeshire) from close to full oxygen saturation at the surface to <5% saturation below ca 10 m. During these summer events, no echinoderms or Ascidia mentula were recorded at depths below 10 - 11 m. Sensitivity assessment. No direct evidence was found on the effect of hypoxia on Novocrania anomala or Protanthea simplex. Based on the evidence of other brachiopods, and anemones, a proportion of the community is likely to be able to survive a short-term anoxic episode. An event at the benchmark level is likely to cause limited mortality among all the characterizing species. Resistance is, therefore recorded as ‘Medium’, resilience as ‘High’ and sensitivity as ‘Low’. | MediumHelp | HighHelp | LowHelp |
Nutrient enrichment [Show more]Nutrient enrichmentBenchmark. Compliance with WFD criteria for good status. Further detail EvidenceJames et al. (1992) conducted a review of brachiopod literature, including brachiopod sensitivity to turbidity and cited Rhodes & Thayer (1991), highlighting the ability of brachiopods to sort and reject particles. The rejected material was removed using mucous streamers, although the extra energy cost of this mechanism was noted. The importance of dissolved organic matter is debated (see James et al., 1992). Steele & Petrovic (1975, 1976, 1979) and Thayer (1986) discussed the importance of the unfused lophophore tentacles in adaptation to highly turbid environments, although whether this makes them better suited to excessive turbidity than bivalves is questioned (Thayer, 1991). Ascidia mentula has been reported in Iberian bays subject to both nutrient-rich upwelling events and anthropogenic pollution (Aneiros et al., 2015). There is some suggestion that there are possible benefits to ascidians from the increased organic content of water; ascidian ‘richness’ in Algeciras Bay was found to increase in higher concentrations of suspended organic matter (Naranjo et al. 1996). Nevertheless, this biotope is considered to be 'Not sensitive' at the pressure benchmark, that assumes compliance with good status as defined by the WFD. | Not relevant (NR)Help | Not relevant (NR)Help | Not sensitiveHelp |
Organic enrichment [Show more]Organic enrichmentBenchmark. A deposit of 100 gC/m2/yr. Further detail EvidenceRhodes & Thayer (1991, cited in James et al.,1992) highlighted the ability of brachiopods to sort and reject particles. The rejected material was removed using mucous streamers, although the extra energy cost of this mechanism was noted. The importance of dissolved organic matter is debated (see James et al., 1992). The lophophore in some brachiopods appears to be able to absorb directly dissolved organic matter from seawater (Storch & Welsch, 1976 cited in Emig, 1997). Steele & Petrovic (1975, 1976, 1979) and Thayer (1986) discussed the importance of the unfused lophophore tentacles in adaptation to highly turbid environments, although whether this makes them better suited to excessive turbidity than bivalves is questioned (Thayer, 1991). Shick (2012) reviewed the uptake of dissolved organic matter in sea anemones and noted (citing Robbins & Shick, 1980) that this feeding mechanism is likely to particularly important for small anemones, which have a proportionally large tentacle surface area. There is some suggestion that there are possible benefits to the ascidians from the increased organic content of water. Ascidian ‘richness’ in Algeciras Bay was found to increase in higher concentrations of suspended organic matter (Naranjo et al. 1996). Kocak & Kucuksezgin (2000) noted that Ciona intestinalis was one of the rapid breeding opportunistic species that tended to be dominant in Turkish harbours enriched by organic pollutants and was frequently found in polluted environments (Carver et al., 2006). Ascidia mentula has been reported in Iberian bays subject to both nutrient-rich upwelling events and anthropogenic organic pollution (Aneiros et al., 2015). Sensitivity assessment. All of the characterizing species are filter feeders and dissolved organic matter is an important food source. However, organic enrichment can lead to eutrophication or hypoxia. But ‘No evidence’ as to the effects of organic enrichment in sublittoral rock was found. | No evidence (NEv)Help | Not relevant (NR)Help | No evidence (NEv)Help |
Physical Pressures
Use [show more] / [show less] to open/close text displayed
| Resistance | Resilience | Sensitivity | |
Physical loss (to land or freshwater habitat) [Show more]Physical loss (to land or freshwater habitat)Benchmark. A permanent loss of existing saline habitat within the site. Further detail EvidenceAll marine habitats and benthic species are considered to have a resistance of ‘None’ to this pressure and to be unable to recover from a permanent loss of habitat (resilience is ‘Very low’). Sensitivity within the direct spatial footprint of this pressure is, therefore ‘High’. Although no specific evidence is described confidence in this assessment is ‘High’, due to the incontrovertible nature of this pressure. | NoneHelp | Very LowHelp | HighHelp |
Physical change (to another seabed type) [Show more]Physical change (to another seabed type)Benchmark. Permanent change from sedimentary or soft rock substrata to hard rock or artificial substrata or vice-versa. Further detail EvidenceThe replacement of rock by sediment would be a fundamentally change the physical character of the biotope, which would be lost. Resistance to the pressure is considered ‘None’, and resilience ‘Very low’ (a permanent change). Sensitivity has been assessed as ‘High’. | NoneHelp | Very LowHelp | HighHelp |
Physical change (to another sediment type) [Show more]Physical change (to another sediment type)Benchmark. Permanent change in one Folk class (based on UK SeaMap simplified classification). Further detail Evidence‘Not relevant’ to biotopes occurring on bedrock. | Not relevant (NR)Help | Not relevant (NR)Help | Not relevant (NR)Help |
Habitat structure changes - removal of substratum (extraction) [Show more]Habitat structure changes - removal of substratum (extraction)Benchmark. The extraction of substratum to 30 cm (where substratum includes sediments and soft rock but excludes hard bedrock). Further detail EvidenceThe species characterizing this biotope are epifauna or epiflora occurring on rock and would be sensitive to the removal of the habitat. However, extraction of rock substratum is considered unlikely and this pressure is considered to be ‘Not relevant’ to hard substratum habitats. | Not relevant (NR)Help | Not relevant (NR)Help | Not relevant (NR)Help |
Abrasion / disturbance of the surface of the substratum or seabed [Show more]Abrasion / disturbance of the surface of the substratum or seabedBenchmark. Damage to surface features (e.g. species and physical structures within the habitat). Further detail EvidenceNovocrania anomala is a sessile brachiopod protected by a calcified shell. However its shell is not massively strong and physical disturbance (such as a passing scallop dredge) will probably cause damage and death (Long & Stricker, 1991; James et al., 1992; Jackson, 2000). Protanthea simplex is delicate and soft-bodied. Abrasion is highly likely to cause mortality. Both Ciona intestinalis and Ascidia mentula are large, emergent, sessile ascidians, and physical disturbance is likely to cause damage and mortality. Emergent epifauna are generally very intolerant of disturbance from fishing gear (Jennings & Kaiser, 1998). However, studies have shown Ascidia spp. to become more abundant following disturbance events (Bradshaw et al., 2000). Sensitivity assessment. Whilst the hard shell of Novocrania anomala would afford some protection from a lighter abrasion event, both the ascidians and the soft-bodied Protanthea simplex are likely to be significantly affected. Resistance is, therefore, assessed as ‘Low’, resilience as ‘Medium’, and sensitivity as ‘Medium’. | LowHelp | MediumHelp | MediumHelp |
Penetration or disturbance of the substratum subsurface [Show more]Penetration or disturbance of the substratum subsurfaceBenchmark. Damage to sub-surface features (e.g. species and physical structures within the habitat). Further detail EvidenceThe species characterizing this biotope group are epifauna or epiflora occurring on rock which is resistant to subsurface penetration. The assessment for abrasion at the surface only is therefore considered to equally represent sensitivity to this pressure. This pressure is thought ‘Not Relevant’ to hard rock biotopes | Not relevant (NR)Help | Not relevant (NR)Help | Not relevant (NR)Help |
Changes in suspended solids (water clarity) [Show more]Changes in suspended solids (water clarity)Benchmark. A change in one rank on the WFD (Water Framework Directive) scale e.g. from clear to intermediate for one year. Further detail EvidenceRhodes and Thayer’s (1991, cited in James et al., 1992) highlighted the ability of brachiopods to sort and reject particles. The rejected material was removed using mucous streamers, although the extra energy cost of this mechanism was noted. The importance of dissolved organic matter as a food source was debated (see James et al., 1992). Steele-Petrović (1975, 1976, 1979) and Thayer (1986) discussed the importance of the unfused lophophore tentacles in adaptation to turbid environments, although whether this makes them better suited to excessive turbidity than bivalves was questioned (Thayer, 1991). Whilst brachiopods appear well suited to cope with turbid conditions, Rudwick (1965, 1970, cited in James et al., 1992) noted that the sessile brachiopods, unable to maintain themselves at the surface of a rapidly accreting substratum, would not tolerate sedimentation. Increased siltation may clog the anemone's tentacles and interfere with feeding. Clearing the sediment will require increased energetic expenditure and loss of condition may result. Ciona intestinalis frequently occurs in habitats close to harbours and marinas with high levels of silt and suspended matter (Carver et al., 2006; Kocak & Kucuksezgin, 2000). Naranjo et al. (1996) described Ciona intestinalis as having a large body and siphons that have wide apertures that helps prevent blocking. Increased suspended sediment may potentially have some detrimental effects in clogging up feeding filtration mechanisms, however, there are possible benefits from increased suspended sediment as ascidian ‘richness’ in Algeciras Bay was found to increase in higher concentrations of suspended organic matter (Naranjo et al. 1996). In high (up to 300 mg/l of inorganic and 2.5 x107 cells/l) suspended particulate concentrations, the active rejection mechanism (squirting) is increased in Ciona intestinalis with no discrimination between organic and inorganic particulates observed in any of the ascidians in the study (Robbins, 1984a). Despite these observations, the turbidity tolerance level for this species is not well established. Robbins (1985a) found that continual exposure to elevated levels of inorganic particulates (>25 mg/l) arrested the growth rate of Ciona intestinalis and exposure to 600 mg/l resulted in 50% mortality after 12-15 days and 100% mortality after 3 weeks. It was suggested that because this species can only “squirt” to clear the branchial sac, it may be vulnerable to clogging under heavy sediment loads. Ascidia mentula has been shown to decrease absolute (instantaneous) rate of pumping in high suspended particulate concentrations, whilst filtration efficiency remained unchanged (Robbins, 1984a). However, specific data on the sensitivity to suspended sediment is lacking. Sensitivity assessment. Whilst increase in turbidity may result in increased energy expenditure for some of the characterizing species, all have methods to cope with increased turbidity. Assuming the biotope typically occurs in intermediate turbidity (10-100 mg/l), an increase at the benchmark level (to medium, 100-300 mg/l) is unlikely to cause significant mortality amongst the characterizing species and resistance is assessed as ‘High’, Resilience as ‘High’ and the biotope is considered to be ‘Not sensitive’. | HighHelp | HighHelp | Not sensitiveHelp |
Smothering and siltation rate changes (light) [Show more]Smothering and siltation rate changes (light)Benchmark. ‘Light’ deposition of up to 5 cm of fine material added to the seabed in a single discrete event. Further detail EvidenceProtanthea simplex is a small (2 cm high), delicate anemone (Jackson, 2008). Smothering with 5 cm of sediment is likely to cause physical damage to the anemone as well as restricting respiration and preventing feeding. In the event of burial, Novocrania anomala is up to 1.5 cm in size (Jackson, 2000), can close their shell and survive in low oxygen concentrations, which can be tolerated for a few days (Thayer, 1981 cited in James et al., 1992). Longer term, smothering by sediment will prevent feeding and result in anoxic conditions, eventually resulting in mortality. Adults are sessile, being permanently cemented to the substratum (Jackson, 2000) and Rudwick (1965, 1970, cited in James et al., 1992) noted that the sessile brachiopods, unable to maintain themselves at the surface of a rapidly accreting substratum, would not tolerate sedimentation. The solitary ascidians considered in this report are permanently attached to the substratum and are active suspension feeders. Smothering with 5 cm of sediment is likely to only affect a small proportion of the population because the adults reach up to 15 cm and 18 cm in length for Ciona intestinalis and Ascidia mentula respectively (Rowley, 2008; Jackson, 2008) and frequently inhabit vertical surfaces (Jackson, 2008), Recovery should be rapid, facilitated by the remaining adults. Sensitivity assessment. Smothering by 5 cm of sediment is likely to impact the characterizing species of this biotope including the sessile ascidians and Novocrania anomala. The biotope often occurs on vertical walls which would protect the characterizing species from sedimentation effects. In flatter areas where sedimentation could occur, given the low water flow rates experienced by these biotopes (Weak to very weak), it is unlikely that the sediment would be removed very rapidly and it is unlikely that Novocrania anomala could survive. Resistance has been assessed as ‘Medium’, resilience as ‘High’ and sensitivity has been assessed as ‘Low’ at the benchmark level. | MediumHelp | HighHelp | LowHelp |
Smothering and siltation rate changes (heavy) [Show more]Smothering and siltation rate changes (heavy)Benchmark. ‘Heavy’ deposition of up to 30 cm of fine material added to the seabed in a single discrete event. Further detail EvidenceProtanthea simplex is a small (2 cm high), delicate anemone (Jackson, 2008). Smothering with 30 cm of sediment is highly likely to cause physical damage to the anemone as well as restricting respiration and preventing feeding. In the event of burial, the dorsal valve of Novocrania anomala is up to 1.5 cm in size (Jackson, 2000), can close their shell and survive in low oxygen concentrations, which can be tolerated for a few days (Thayer, 1981 cited in James et al., 1992). Longer term, smothering by sediment will prevent feeding and result in anoxic conditions, eventually resulting in mortality. Adults are sessile, being permanently cemented to the substratum (Jackson, 2000) and Rudwick (1965, 1970, cited in James et al., 1992) noted that the sessile brachiopods, unable to maintain themselves at the surface of a rapidly accreting substratum, would not tolerate sedimentation. The solitary ascidians considered in this report are permanently attached to the substratum and are active suspension feeders. Smothering with 30 cm of sediment is likely to affect a proportion of the population because, although the adults reach up to 15 cm and 18 cm in length for Ciona intestinalis and Ascidia mentula respectively (Rowley, 2008; Jackson, 2008), they frequently inhabit vertical surfaces (Jackson, 2008). Recovery should be rapid, facilitated by the remaining adults. Sensitivity assessment. Smothering by 30 cm of sediment is likely to impact the characterizing species of this biotope especially the sessile ascidians and Novocrania anomala. The biotope often occurs on vertical walls which would protect the characterizing species from sedimentation effects. In flatter areas where sedimentation could occur, given the low water flow rates experienced by these biotopes (weak to very weak), it is unlikely that the sediment would be removed very rapidly and few Novocrania anomala and ascidians would survive. The resilience assessment assumes removal of the sediment, however, depending on the conditions, this may not be possible. Resistance has been assessed as ‘Low’, resilience as ‘Medium’ and sensitivity has been assessed as ‘Medium’ at the benchmark level. | LowHelp | MediumHelp | MediumHelp |
Litter [Show more]LitterBenchmark. The introduction of man-made objects able to cause physical harm (surface, water column, seafloor or strandline). Further detail EvidenceNot assessed | Not Assessed (NA)Help | Not assessed (NA)Help | Not assessed (NA)Help |
Electromagnetic changes [Show more]Electromagnetic changesBenchmark. A local electric field of 1 V/m or a local magnetic field of 10 µT. Further detail EvidenceNo evidence | No evidence (NEv)Help | Not relevant (NR)Help | No evidence (NEv)Help |
Underwater noise changes [Show more]Underwater noise changesBenchmark. MSFD indicator levels (SEL or peak SPL) exceeded for 20% of days in a calendar year. Further detail EvidenceNo evidence could be found for the effects of noise or vibrations on Novocrania anomala or Protanthea simplex but they are considered unlikely to be sensitive to this pressure. McDonald (2014) studied the effect of generator noise on fouling of four vessels by Ciona intestinalis and found that fouling was highest at locations closest to the generators and lowest furthest away from the generators. Subsequent in vitro experiments demonstrated that larvae settled much faster in the presence of noise (2 h- 20 h compared with 6 h-26 h for control), underwent metamorphosis more rapidly (between 10 and 20 h compared with ca 22 h) and had a markedly increased survival rate to maturity (90-100% compared with 66%). Other studies also reported that noise emissions from vessels promoted fouling by organisms including ascidians (Stanley et al., 2016). Sensitivity assessment: Resistance to this pressure is assessed as 'High' and resilience as 'High'. This biotope is therefore considered to be 'Not sensitive'. | Not relevant (NR)Help | Not relevant (NR)Help | Not relevant (NR)Help |
Introduction of light or shading [Show more]Introduction of light or shadingBenchmark. A change in incident light via anthropogenic means. Further detail EvidenceWhilst most brachiopods are believed to detect light, the means of detection has been questioned and few studies have been undertaken (James et al., 1992). It should be noted that McCammon (1973) reported that Magellania venosa was not sensitive to light. Anemones have been reported to respond to light (Parker, 1919 cited in Sebens, 1981). The ascidian Dendrodoa spp. larvae do not possess an eye spot, and light does not appear to affect locomotion (Mackie & Bone, 1976). Sensitivity assessment. Whilst there is evidence that some characterizing species respond to light, either as larvae or as adults, change at the benchmark level is unlikely to cause mortality among any of the species assessed, and resistance and resilience are both recorded as ‘High’, and the biotope is considered ‘Not sensitive’ to this pressure. | HighHelp | HighHelp | Not sensitiveHelp |
Barrier to species movement [Show more]Barrier to species movementBenchmark. A permanent or temporary barrier to species movement over ≥50% of water body width or a 10% change in tidal excursion. Further detail EvidenceNot relevant, barriers and changes in tidal excursion are not relevant to biotopes restricted to open waters. | Not relevant (NR)Help | Not relevant (NR)Help | Not relevant (NR)Help |
Death or injury by collision [Show more]Death or injury by collisionBenchmark. Injury or mortality from collisions of biota with both static or moving structures due to 0.1% of tidal volume on an average tide, passing through an artificial structure. Further detail EvidenceNot relevant to seabed habitats. NB. Collision by grounding vessels is addressed under ‘surface abrasion’. | Not relevant (NR)Help | Not relevant (NR)Help | Not relevant (NR)Help |
Visual disturbance [Show more]Visual disturbanceBenchmark. The daily duration of transient visual cues exceeds 10% of the period of site occupancy by the feature. Further detail EvidenceNot relevant | Not relevant (NR)Help | Not relevant (NR)Help | Not relevant (NR)Help |
Biological Pressures
Use [show more] / [show less] to open/close text displayed
| Resistance | Resilience | Sensitivity | |
Genetic modification & translocation of indigenous species [Show more]Genetic modification & translocation of indigenous speciesBenchmark. Translocation of indigenous species or the introduction of genetically modified or genetically different populations of indigenous species that may result in changes in the genetic structure of local populations, hybridization, or change in community structure. Further detail EvidenceCiona intestinalis is considered a fouling species and adheres readily to the hulls of vessels, and is considered an invasive species in the USA, Chile, Western Australia, New Zealand, Canada and South Africa (Millar 1966; McDonald 2004; Blum et al. 2007; Ramsay et al. 2008, 2009; Dumont et al.,2011). Whilst there have been novel proposals to farm Ciona intestinalis as a potential feedstock for aquaculture in Sweden (Laupsa, 2015), there is no evidence to suggest such farming exists. No evidence for Novocrania anomala or Protanthea simplex could be found. Therefore, there is currently ‘No evidence’ on which to assess this pressure. | No evidence (NEv)Help | Not relevant (NR)Help | No evidence (NEv)Help |
Introduction of microbial pathogens [Show more]Introduction of microbial pathogensBenchmark. The introduction of relevant microbial pathogens or metazoan disease vectors to an area where they are currently not present (e.g. Martelia refringens and Bonamia, Avian influenza virus, viral Haemorrhagic Septicaemia virus). Further detail EvidenceLittle is known about brachiopod disease, therefore there is 'No evidence' on which to base an assessment. However, James et al. (1992) highlighted significant shell and soft tissue regenerative capabilities. McCammon (1973) recorded infection of the lophophore tentacles of Magellania venosa by an unidentified protist which destroyed connective tissues and ultimately resulted in death. McCammon (1972) also observed brachiopods subject to stress being prone to fungal infection which exhibited black spots forming on the shell. More generally, James et al. (1992) discussed brachiopod success in aquaria and suggests that they are relatively hardy. No evidence for diseases affecting Protanthea simplex could be found. There appears to be little research into ascidian diseases, particularly in the Atlantic. The parasite Lankesteria ascidiae targets the digestive tubes and can cause ‘long faeces syndrome’ in Ciona intestinalis (although it has also been recorded in other species). Mortality occurs in severely affected individuals within about a week following first symptoms. (Mita et al., 2012). There is ‘No evidence’ to support an assessment of this pressure. | No evidence (NEv)Help | Not relevant (NR)Help | No evidence (NEv)Help |
Removal of target species [Show more]Removal of target speciesBenchmark. Removal of species targeted by fishery, shellfishery or harvesting at a commercial or recreational scale. Further detail EvidenceAmong brachiopods, only the lingulids (Lingula spp.) have been fished commercially, and on a very small scale (Printrakoon & Kamlung-ek, 2013). No evidence for brachiopod extraction in the British Isles was found. Whilst unlikely, should targeted extraction occur, it is likely to have a significant impact on the characterizing species. Whilst anemones are not sessile, they are very slow moving and are therefore likely to be as exposed to extraction as the sessile Novocrania anomala and ascidians. 'Not relevant' as none of the characterizing species are targeted. | LowHelp | MediumHelp | MediumHelp |
Removal of non-target species [Show more]Removal of non-target speciesBenchmark. Removal of features or incidental non-targeted catch (by-catch) through targeted fishery, shellfishery or harvesting at a commercial or recreational scale. Further detail EvidenceThe characteristic species probably compete for space within the biotope, so that loss of one species would probably have little if any effect on the other members of the community. Removal of the characteristic epifauna due to by-catch is likely to remove a proportion of the biotope and change the biological character of the biotope. These direct, physical impacts are assessed through the abrasion and penetration of the seabed pressures. The sensitivity assessment for this pressure considers any biological/ecological effects resulting from the removal of non-target species on this biotope. Therefore, resistance is assessed as ‘Low’, resilience as ‘Medium’ and sensitivity assessed as ‘Medium’. This biotope may be removed or damaged by static or mobile gears that are targeting other species (see physical pressures above). | LowHelp | MediumHelp | MediumHelp |
Introduction or spread of invasive non-indigenous species (INIS) Pressures
Use [show more] / [show less] to open/close text displayed
| Resistance | Resilience | Sensitivity | |
Other INIS [Show more]Other INISEvidenceThis biotope is classified as circalittoral and therefore no algal species have been considered. Crepidula fornicata larvae require hard substrata for settlement. It prefers muddy gravelly, shell-rich, substrata that include gravel, or shells of other Crepidula, or other species e.g., oysters, and mussels. It is highly gregarious and seeks out adult shells for settlement, forming characteristic ‘stacks’ of adults. But it also recorded from rock, artificial substrata, and Sabellaria alveolata reefs (Blanchard, 1997, 2009; Bohn et al., 2012, 2013a, 2013b, 2015; De Montaudouin et al., 2018; Hinz et al., 2011; Helmer et al., 2019; Powell-Jennings & Calloway, 2018; Preston et al., 2020; Tillin et al., 2020). Close examination of the literature (2023) shows that evidence of its colonization and density on bedrock in the infralittoral or circalittoral was lacking. Tillin et al. (2020) suggested that Crepidula could colonize circalittoral rock due to its presence on tide-swept rough grounds in the English Channel (Hinz et al., 2011). However, Hinz et al. (2011) reported that Crepidula fornicata only dominated one assemblage (with an average of 181 individuals per trawl) on gravel substratum with boulders. Bohn et al. (2015) noted that Crepidula occurred at low density or was absent in areas dominated by boulders, and Bohn et al. (2013a, 2013b, 2015) and Preston et al. (2020) showed that while Crepidula could settle on slate panels or ‘stone’ it preferred shell, especially that of conspecifics. In addition, no evidence was found of the effect of Crepidula populations on faunal turf-dominated habitats. It was only recorded at low density (0.1-0.9/m2) in one faunal turf biotope (CR.MCR.CFaVS.CuSpH.As) (JNCC, 2015). Faunal turfs are dominated by suspension feeders so larval predation is probably high, which may prevent colonization by Crepidula. Also, faunal turf species actively compete for space and many are fast growing and opportunistic, so may out-compete Crepidula for space even if it gained a foothold in the community. Didemnum vexillum is an invasive colonial sea squirt native to Asia which was first recorded in the UK in Darthaven Marina, Dartmouth in 2005. Didemnum vexillum can form extensive mats over the substrata it colonizes, binding boulders and cobbles and altering the host habitat (Griffith et al., 2009). Didemnum vexillum can also grow over and smother the resident biological community. Recent surveys within Holyhead Marina, North Wales have found Didemnum vexillum growing on and smothering native tunicate communities, including Ciona intestinalis (Griffith et al., 2009). Due to the rapid-re-colonization of Didemnum vexillum eradication attempts have to date failed. Presently Didemnum vexillum is isolated to several sheltered locations in the UK (NBN, 2015), however, Didemnum vexillum has successfully colonized offshore in Georges Bank, USA (Lengyel et al., 2009). Styela clava, another INIS ascidian, was first recorded in the UK at Plymouth in 1952 (Eno et al., 1997). Sensitivity assessment. The circalittoral rock characterizing this biotope could be suitable for colonization by Crepidula fornicata due to the sheltered to extremely sheltered wave conditions, where water is mediated by tidal flow rather than wave actions. Crepidula has been recorded from the lower intertidal to ca 160 m in depth but is most common in the shallow subtidal above 50 m (Blanchard, 1997; Thieltges et al., 2003; Bohn et al., 2012, 2015; Hinz et al., 2011; OBIS, 2023; Tillin et al., 2020), therefore, colonization of Crepidula would be limited to low densities in deeper examples of the biotope. In addition, no evidence was found of the effect of Crepidula populations on faunal turf-dominated habitats or infralittoral or circalittoral rock habitats. Didemnum vexillum could pose a potential threat. At present, there is 'Insufficient evidence' to suggest that the circalittoral rock biotopes are sensitive to colonization by Crepidula fornicata or other invasive species; further evidence is required. | Insufficient evidence (IEv)Help | Not relevant (NR)Help | Help |
Bibliography
Álvarez, F. & Emig, C., 2000. Brachiopoda from the Luso-Iberian zone. I. Biology and ecology. In The Millennium Brachiopod Congress, London, 2000. Abstracts.
Aneiros, F., Rubal, M., Troncoso, J.S. & Bañón, R., 2015. Subtidal benthic megafauna in a productive and highly urbanised semi-enclosed bay (Ría de Vigo, NW Iberian Peninsula). Continental Shelf Research, 110, 16-24.
Bay-Nouailhat, W., 2007. Description de Sarcodictyon roseum. [cited 17.03.2016]. Available from: http://www.mer-littoral.org/05/sarcodictyon-roseum.php
Blanchard, M., 2009. Recent expansion of the slipper limpet population (Crepidula fornicata) in the Bay of Mont-Saint-Michel (Western Channel, France). Aquatic Living Resources, 22 (1), 11-19. DOI https://doi.org/10.1051/alr/2009004
Blanchard, M., 1997. Spread of the slipper limpet Crepidula fornicata (L.1758) in Europe. Current state and consequences. Scientia Marina, 61, Supplement 9, 109-118. Available from: http://scimar.icm.csic.es/scimar/index.php/secId/6/IdArt/290/
Blum, J.C., Chang, A.L., Liljesthröm, M., Schenk, M.E., Steinberg, M.K. & Ruiz, G.M., 2007. The non-native solitary ascidian Ciona intestinalis (L.) depresses species richness. Journal of Experimental Marine Biology and Ecology, 342 (1), 5-14.
Bohn, K., Richardson, C. & Jenkins, S., 2012. The invasive gastropod Crepidula fornicata: reproduction and recruitment in the intertidal at its northernmost range in Wales, UK, and implications for its secondary spread. Marine Biology, 159 (9), 2091-2103. DOI https://doi.org/10.1007/s00227-012-1997-3
Bohn, K., Richardson, C.A. & Jenkins, S.R., 2015. The distribution of the invasive non-native gastropod Crepidula fornicata in the Milford Haven Waterway, its northernmost population along the west coast of Britain. Helgoland Marine Research, 69 (4), 313.
Bohn, K., Richardson, C.A. & Jenkins, S.R., 2013a. Larval microhabitat associations of the non-native gastropod Crepidula fornicata and effects on recruitment success in the intertidal zone. Journal of Experimental Marine Biology and Ecology, 448, 289-297. DOI https://doi.org/10.1016/j.jembe.2013.07.020
Bohn, K., Richardson, C.A. & Jenkins, S.R., 2013b. The importance of larval supply, larval habitat selection and post-settlement mortality in determining intertidal adult abundance of the invasive gastropod Crepidula fornicata. Journal of Experimental Marine Biology and Ecology, 440, 132-140. DOI https://doi.org/10.1016/j.jembe.2012.12.008
Bradshaw, C., Veale, L.O., Hill, A.S. & Brand, A.R., 2000. The effects of scallop dredging on gravelly seabed communities. In: Effects of fishing on non-target species and habitats (ed. M.J. Kaiser & de S.J. Groot), pp. 83-104. Oxford: Blackwell Science.
Butman, C.A., 1987. Larval settlement of soft-sediment invertebrates: the spatial scales of pattern explained by active habitat selection and the emerging role of hydrodynamical processes. Oceanography and Marine Biology: an Annual Review, 25, 113-165.
Caputi, L., Crocetta, F., Toscano, F., Sordino, P. & Cirino, P., 2015. Long-term demographic and reproductive trends in Ciona intestinalis sp. A. Marine Ecology, 36 (1), 118-128.
Carlgren, O., 1921. Actiniaria. Pt. 1. Danish Ingolf Expedition, Vol. V, No. 9., pp. 31. Copenhagen: Bianco Luno.
Carlgren, O., 1949. A survey of the Ptychodactiaria, Corallimorpharia and Actiniaria. Kungliga Svenska Vetenskapsakadamiens Handlingar, Series 4, 1, 16-110.
Carver, C., Mallet, A. & Vercaemer, B., 2006. Biological synopsis of the solitary tunicate Ciona intestinalis. Canadian Manuscript Report of Fisheries and Aquatic Science, No. 2746, v + 55 p. Bedford Institute of Oceanography, Dartmouth, Nova Scotia.
Castège, I., Milon, E. & Pautrizel, F., 2014. Response of benthic macrofauna to an oil pollution: Lessons from the “Prestige” oil spill on the rocky shore of Guéthary (south of the Bay of Biscay, France). Deep Sea Research Part II: Topical Studies in Oceanography, 106, 192-197.
Chia, F.S. & Spaulding, J.G., 1972. Development and juvenile growth of the sea anemone Tealia crassicornis. Biological Bulletin, Marine Biological Laboratory, Woods Hole, 142, 206-218.
Cole, S., Codling, I.D., Parr, W. & Zabel, T., 1999. Guidelines for managing water quality impacts within UK European Marine sites. Natura 2000 report prepared for the UK Marine SACs Project. 441 pp., Swindon: Water Research Council on behalf of EN, SNH, CCW, JNCC, SAMS and EHS. [UK Marine SACs Project.]. Available from: http://ukmpa.marinebiodiversity.org/uk_sacs/pdfs/water_quality.pdf
Connor, D.W., Allen, J.H., Golding, N., Howell, K.L., Lieberknecht, L.M., Northen, K.O. & Reker, J.B., 2004. The Marine Habitat Classification for Britain and Ireland. Version 04.05. ISBN 1 861 07561 8. In JNCC (2015), The Marine Habitat Classification for Britain and Ireland Version 15.03. [2019-07-24]. Joint Nature Conservation Committee, Peterborough. Available from https://mhc.jncc.gov.uk/
Connor, D.W., Dalkin, M.J., Hill, T.O., Holt, R.H.F. & Sanderson, W.G., 1997a. Marine biotope classification for Britain and Ireland. Vol. 2. Sublittoral biotopes. Joint Nature Conservation Committee, Peterborough, JNCC Report no. 230, Version 97.06., Joint Nature Conservation Committee, Peterborough, JNCC Report no. 230, Version 97.06.
Davies, C.E. & Moss, D., 1998. European Union Nature Information System (EUNIS) Habitat Classification. Report to European Topic Centre on Nature Conservation from the Institute of Terrestrial Ecology, Monks Wood, Cambridgeshire. [Final draft with further revisions to marine habitats.], Brussels: European Environment Agency.
De Montaudouin, X., Blanchet, H. & Hippert, B., 2018. Relationship between the invasive slipper limpet Crepidula fornicata and benthic megafauna structure and diversity, in Arcachon Bay. Journal of the Marine Biological Association of the United Kingdom, 98 (8), 2017-2028. DOI https://doi.org/10.1017/s0025315417001655
Diaz, R.J. & Rosenberg, R., 1995. Marine benthic hypoxia: a review of its ecological effects and the behavioural responses of benthic macrofauna. Oceanography and Marine Biology: an Annual Review, 33, 245-303.
Dumont, C., Gaymer, C. & Thiel, M., 2011. Predation contributes to invasion resistance of benthic communities against the non-indigenous tunicate Ciona intestinalis. Biological Invasions, 13 (9), 2023-2034.
Dybern, B.I., 1965. The life cycle of Ciona intestinalis (L.) f. typica in relation to the environmental temperature. Oikos, 16, 109-131.
Dybern, B.I., 1967. The distribution and salinity tolerance of Ciona intestinalis (L.) f. typica with special reference to the waters around southern Scandinavia. Ophelia, 4 (2), 207-226.
Dybern, B.I., 1969. Distribution and ecology of ascidians in Kviturdvikpollen and Vågsböpollen on the west coast of Norway. Sarsia, 37 (1), 21-40.
Ellington, W.R., 1982. Metabolic responses of the sea anemone Bunodosoma cavernata (Bosc) to declining oxygen tensions and anoxia. Physiological zoology, 240-249.
Emig, C.C., 1997. Ecology of inarticulated brachiopods. In Treatise of Invertebrate Paleontology (Kaesler, RL; editor). Part H, 473-495. Geological Society of America and the University of Kansas Press.
Eno, N.C., Clark, R.A. & Sanderson, W.G. (ed.) 1997. Non-native marine species in British waters: a review and directory. Peterborough: Joint Nature Conservation Committee.
Fent, K., 1996. Ecotoxicology of organotin compounds. Critical reviews in toxicology, 26 (1), 3-117.
Glantz, M.H., 2005. Climate variability, climate change and fisheries. Cambridge: Cambridge University Press.
Griffith, K., Mowat, S., Holt, R.H., Ramsay, K., Bishop, J.D., Lambert, G. & Jenkins, S.R., 2009. First records in Great Britain of the invasive colonial ascidian Didemnum vexillum Kott, 2002. Aquatic Invasions, 4 (4), 581-590.
Gulliksen, B., 1977. Studies from the “UWL Helgoland” on the macrobenthic fauna of rocks and boulders in Lübeck Bay (western Baltic Sea). Helgolander Wissenschaftliche Meeresuntersuchungen, 30(1-4), 519-526.
Hammond, L., 1983. Experimental studies of salinity tolerance, burrowing behavior and pedicle regeneration in Lingula anatina (Brachiopoda, Inarticulata). Journal of Paleontology, 1311-1316.
Hartnoll, R.G., 1998. Circalittoral faunal turf biotopes: an overview of dynamics and sensitivity characteristics for conservation management of marine SACs, Volume VIII. Scottish Association of Marine Sciences, Oban, Scotland, 109 pp. [UK Marine SAC Project. Natura 2000 reports.] Available from: http://ukmpa.marinebiodiversity.org/uk_sacs/pdfs/circfaun.pdf
Havenhand, J. & Svane, I., 1989. Larval behaviour, recruitment, and the role of adult attraction in Ascidia mentula O. F. Mueller: Reproduction, genetics and distributions of marine organisms. 23rd European Marine Biology Symposium. Olsen and Olsen, 127-132.
Havenhand, J.N. & Svane, I., 1991. Roles of hydrodynamics and larval behaviour in determining spatial aggregation in the tunicate Ciona intestinalis. Marine Ecology Progress Series, 68, 271-276.
Hayward, P.J. & Ryland, J.S. (ed.), 1995. The marine fauna of the British Isles and north-west Europe. Volume 2. Molluscs to Chordates. Oxford Science Publications. Oxford: Clarendon Press.
Heip, C.H., Keegan, B.F. & Lewis, J.R., 1985. Long-Term Changes in Coastal Benthic Communities. In Proceedings of a Symposium, held in Brussels, Belgium, December 9–12, 1985: Springer Science & Business Media.
Helmer, L., Farrell, P., Hendy, I., Harding, S., Robertson, M. & Preston, J., 2019. Active management is required to turn the tide for depleted Ostrea edulis stocks from the effects of overfishing, disease and invasive species. Peerj, 7 (2). DOI https://doi.org/10.7717/peerj.6431
Herreid, C.F., 1980. Hypoxia in invertebrates. Comparative Biochemistry and Physiology Part A: Physiology, 67 (3), 311-320. DOI https://doi.org/10.1016/S0300-9629(80)80002-8
Hinz, H., Capasso, E., Lilley, M., Frost, M. & Jenkins, S.R., 2011. Temporal differences across a bio-geographical boundary reveal slow response of sub-littoral benthos to climate change. Marine Ecology Progress Series, 423, 69-82. DOI https://doi.org/10.3354/meps08963
Hiscock, K. & Hoare, R., 1975. The ecology of sublittoral communities at Abereiddy Quarry, Pembrokeshire. Journal of the Marine Biological Association of the United Kingdom, 55 (4), 833-864.
Hiscock, K., 1983. Water movement. In Sublittoral ecology. The ecology of shallow sublittoral benthos (ed. R. Earll & D.G. Erwin), pp. 58-96. Oxford: Clarendon Press.
Hiscock, K., 1985. Littoral and sublittoral monitoring in the Isles of Scilly. September 22nd to 29th, 1984. Nature Conservancy Council, Peterborough, CSD Report, no. 562., Field Studies Council Oil Pollution Research Unit, Pembroke.
Holt, T.J., Jones, D.R., Hawkins, S.J. & Hartnoll, R.G., 1995. The sensitivity of marine communities to man induced change - a scoping report. Countryside Council for Wales, Bangor, Contract Science Report, no. 65.
Jackson, A. 2000. Novocrania anomala, A brachiopod. In Tyler-Walters H. and Hiscock K. (eds) Marine Life Information Network: Biology and Sensitivity Key Information Reviews, [on-line]. Plymouth: Marine Biological Association of the United Kingdom. [17.03.16] Available from: http://www.marlin.ac.uk/species/detail/1331
Jackson, A., 2008. Ciona intestinalis. A sea squirt. Marine Life Information Network: Biology and Sensitivity Key Information Sub-programme [On-line]. Plymouth: Marine Biological Association of the United Kingdom. [cited 16/12/15] Available from:
Jackson, A. 2008b. Protanthea simplex, Sealoch anemone. In Tyler-Walters H. and Hiscock K. (eds) Marine Life Information Network: Biology and Sensitivity Key Information Reviews, [on-line]. Plymouth: Marine Biological Association of the United Kingdom. [17.03.2016] Available from: http://www.marlin.ac.uk/species/detail/1345
James, M.A., Ansell, A.D., Collins, M.J., Curry, G.B., Peck, L.S. & Rhoda, M.C., 1992. Biology of living brachiopods. Advances in Marine Biology, 28, 175-387.
Jennings, S. & Kaiser, M.J., 1998. The effects of fishing on marine ecosystems. Advances in Marine Biology, 34, 201-352.
JNCC (Joint Nature Conservation Committee), 2022. The Marine Habitat Classification for Britain and Ireland Version 22.04. [Date accessed]. Available from: https://mhc.jncc.gov.uk/
JNCC (Joint Nature Conservation Committee), 1999. Marine Environment Resource Mapping And Information Database (MERMAID): Marine Nature Conservation Review Survey Database. [on-line] http://www.jncc.gov.uk/mermaid
Kerby, J.L., Richards‐Hrdlicka, K.L., Storfer, A. & Skelly, D.K., 2010. An examination of amphibian sensitivity to environmental contaminants: are amphibians poor canaries? Ecology Letters, 13 (1), 60-67.
Kocak, F. & Kucuksezgin, F., 2000. Sessile fouling organisms and environmental parameters in the marinas of the Turkish Aegean coast. Indian journal of marine sciences, 29 (2), 149-157.
Lambert, C.C. & Lambert, G., 1998. Non-indigenous ascidians in southern California harbors and marinas. Marine Biology, 130 (4), 675-688.
Laupsa, M., 2015. Spawning, settlement and growth of Ciona intestinalis in Øygarden, Hardangerfjorden and Kvitsøy. Master's thesis. University of Bergen.
Lengyel, N.L., Collie, J.S. & Valentine, P.C., 2009. The invasive colonial ascidian Didemnum vexillum on Georges Bank - Ecological effects and genetic identification. Aquatic Invasions, 4(1), 143-152. DOI https://doi.org/10.3391/ai.2009.4.1.15
Long, J.A. & Stricker, S.A., 1991. Brachiopoda. In Reproduction of marine invertebrates, Vol. VI. Echinoderms and Lophophorates. (ed. A.C. Giese, J.S. Pearse & V.B. Pearse). California: The Boxwood Press.
Mackie, G. & Bone, Q., 1976. Skin impulses and locomotion in an ascidian tadpole. Journal of the Marine Biological Association of the United Kingdom, 56 (03), 751-768.
Magorrian, B.H. & Service, M., 1998. Analysis of underwater visual data to identify the impact of physical disturbance on horse mussel (Modiolus modiolus) beds. Marine Pollution Bulletin, 36 (5), 354-359. DOI https://doi.org/10.1016/s0025-326x(97)00192-6
Manuel, R.L., 1988. British Anthozoa. Synopses of the British Fauna (New Series) (ed. D.M. Kermack & R.S.K. Barnes). The Linnean Society of London [Synopses of the British Fauna No. 18.]. DOI https://doi.org/10.1002/iroh.19810660505
Marin, M.G., Bresan, M., Beghi, L. & Brunetti, R., 1987. Thermo-haline tolerance of Ciona intestinalis (L. 1767) at different developmental stages. Cahiers de Biologie Marine, 28, 45-57.
Mazouni, N., Gaertner, J. & Deslous-Paoli, J.-M., 2001. Composition of biofouling communities on suspended oyster cultures: an in situ study of their interactions with the water column. Marine Ecology Progress Series, 214, 93-102.
MBA (Marine Biological Association), 1957. Plymouth Marine Fauna. Plymouth: Marine Biological Association of the United Kingdom.
McCammon, H.M., 1972. Establishing and Maintaining Articulate Brachiopods in Aquaria. Journal of Geological Education, 20 (3), 139-142.
McCammon, H.M., 1973. The ecology of Magellania venosa, an articulate brachiopod. Journal of Paleontology, 266-278.
McDonald, J., 2004. The invasive pest species Ciona intestinalis (Linnaeus, 1767) reported in a harbour in southern Western Australia. Marine Pollution Bulletin, 49 (9), 868-870.
Meadows, P.S. & Campbell, J.I., 1972. Habitat selection by aquatic invertebrates. Advances in Marine Biology, 10, 271-382.
Mercier, A., Pelletier, E. & Hamel, J.-F., 1996. Toxicological response of the symbiotic sea anemone Aiptasia pallida to butyltin contamination. Marine Ecology Progress Series, 14 (1), 133-146.
Millar, R., 1971. The biology of ascidians. Advances in marine biology, 9, 1-100.
Millar, R.H., 1966. Tunicata Ascidiacea. Oslo, Universitetsforlaget.
Mita, K., Kawai, N., Rueckert, S. & Sasakura, Y., 2012. Large-scale infection of the ascidian Ciona intestinalis by the gregarine Lankesteria ascidiae in an inland culture system. Diseases of aquatic organisms, 101 (3), 185-195.
Monniot, C. & Monniot, F., 1994. Additions to the inventory of eastern tropical Atlantic ascidians; arrival of cosmopolitan species. Bulletin of Marine Science, 54 (1), 71-93.
Naranjo, S.A., Carballo, J.L., & Garcia-Gomez, J.C., 1996. Effects of environmental stress on ascidian populations in Algeciras Bay (southern Spain). Possible marine bioindicators? Marine Ecology Progress Series, 144 (1), 119-131.
Naylor. P., 2011. Great British Marine Animals, 3rd Edition. Plymouth. Sound Diving Publications.
NBN, 2015. National Biodiversity Network 2015(20/05/2015). https://data.nbn.org.uk/
Nielsen, C., 1991. The development of the brachiopod Crania (Neocrania) anomala (OF Müller) and its phylogenetic significance. Acta Zoologica, 72 (1), 7-28.
Nomaguchi, T.A., Nishijima, C., Minowa, S., Hashimoto, M., Haraguchi, C., Amemiya, S. & Fujisawa, H., 1997. Embryonic thermosensitivity of the ascidian, Ciona savignyi. Zoological Science, 14 (3), 511-515.
Nyholm, K-G., 1959. On the development of the primitive actinian Protanthea simplex, Carlgren. Zoologiska Bidrag Fran Uppsala, Band 33 1958-1962, 69-78.
OBIS (Ocean Biodiversity Information System), 2025. Global map of species distribution using gridded data. Available from: Ocean Biogeographic Information System. www.iobis.org. Accessed: 2025-08-05
Pérès, J.M., 1943. Recherches sur le sang et les organes neuraux des Tuniciers. Annales de l’Institut Oceanographique (Monaco), 21, 229-359.
Petersen, J. & Riisgård, H.U., 1992. Filtration capacity of the ascidian Ciona intestinalis and its grazing impact in a shallow fjord. Marine Ecology-Progress Series, 88, 9-17.
Picton, B.E. & Morrow, C.C., 2015. Ascidia mentula O F Müller, 1776. In Encyclopedia of Marine Life of Britain and Ireland. [cited 26/01/16]. Available from: http://www.habitas.org.uk/marinelife/species.asp?item=ZD1500
Powell-Jennings, C. & Callaway, R., 2018. The invasive, non-native slipper limpet Crepidula fornicata is poorly adapted to sediment burial. Marine Pollution Bulletin, 130, 95-104. DOI https://doi.org/10.1016/j.marpolbul.2018.03.006
Preston, J., Fabra, M., Helmer, L., Johnson, E., Harris-Scott, E. & Hendy, I.W., 2020. Interactions of larval dynamics and substrate preference have ecological significance for benthic biodiversity and Ostrea edulis Linnaeus, 1758 in the presence of Crepidula fornicata. Aquatic Conservation: Marine and Freshwater Ecosystems, 30 (11), 2133-2149. DOI https://doi.org/10.1002/aqc.3446
Prestrud, P., Strøm, H. & Goldman, H.V., 2004. A catalogue of the terrestrial and marine animals of Svalbard. Norsk Polarinstitutt.
Printrakoon, C. & Kamlung-ek, A., 2013. Socioeconomic study and economic value of living fossil, Lingula sp. in mangrove ecosystem in Trat Province, Thailand. Chinese Journal of Population Resources and Environment, 11 (3), 187-199.
Radolović, M., Bakran-Petricioli, T., Petricioli, D., Surić, M. & Perica, D., 2015. Biological response to geochemical and hydrological processes in a shallow submarine cave. Mediterranean Marine Science, 16 (2), 305-324.
Ramsay, A., Davidson, J., Bourque, D. & Stryhn, H., 2009. Recruitment patterns and population development of the invasive ascidian Ciona intestinalis in Prince Edward Island, Canada. Aquatic Invasions, 4 (1), 169-176.
Ramsay, A., Davidson, J., Landry, T. & Stryhn, H., 2008. The effect of mussel seed density on tunicate settlement and growth for the cultured mussel, Mytilus edulis. Aquaculture, 275 (1), 194-200.
Renborg, E., Johannesson, K. & Havenhand, J., 2014. Variable salinity tolerance in ascidian larvae is primarily a plastic response to the parental environment. Evolutionary ecology, 28 (3), 561-572
Riisgård, H.U., Jürgensen, C. & Clausen, T., 1996. Filter-feeding ascidians (Ciona intestinalis) in a shallow cove: implications of hydrodynamics for grazing impact. Journal of Sea Research, 35 (4), 293-300.
Robbins, I., 1984a. The regulation of ingestion rate, at high suspended particulate concentrations, by some phleobranchiate ascidians. Journal of Experimental Marine Biology and Ecology, 82 (1), 1-10.
Rosenberg, R., Hellman, B. & Johansson, B., 1991. Hypoxic tolerance of marine benthic fauna. Marine Ecology Progress Series, 79, 127-131. DOI https://dx.doi.org/10.3354/meps079127
Rowley, S.J., 2008. A sea squirt (Ascidia mentula). Tyler-Walters, H. and Hiscock, K. (eds). Marine Life Information Network: Biology and Sensitivity Key Information Reviews [on-line]. Plymouth: Marine Biological Association of the United Kingdom. [cited 26/01/16]. Available from: http://www.marlin.ac.uk/species/detail/8
Rudwick, M.J.S., 1970. Living and fossil brachiopods. London: Hutchinson University Library
Ruppert, E.E. & Barnes, R.D., 1994. Invertebrate zoology (6th ed.). Fort Worth, USA: Saunders College Publishing.
Sabbadin, A., 1957. Il ciclo biologico di Ciona intestinalis (L.), Molgula manhattensis (De Kay) e Styela plicata (Lesueur) nella Laguna Veneta.
Scheltema, R.S., 1974. Biological interactions determining larval settlement of marine invertebrates. Thalassia Jugoslavica, 10, 263-296.
Sebens, K.P., 1981. Recruitment in a Sea Anemone Population: Juvenile Substrate Becomes Adult Prey. Science, 213 (4509), 785-787.
Sebens, K.P., 1985. Community ecology of vertical rock walls in the Gulf of Maine: small-scale processes and alternative community states. In The Ecology of Rocky Coasts: essays presented to J.R. Lewis, D.Sc. (ed. P.G. Moore & R. Seed), pp. 346-371. London: Hodder & Stoughton Ltd.
Sebens, K.P., 1986. Spatial relationships among encrusting marine organisms in the New England subtidal zone. Ecological Monographs, 56, 73-96. DOI https://doi.org/10.2307/2937271
Seeley, R., 2006. Sealife survey. MarLIN news, 9.
Service, M. & Magorrian, B.H., 1997. The extent and temporal variation of disturbance to epibenthic communities in Strangford Lough, Northern Ireland. Journal of the Marine Biological Association of the United Kingdom, 77, 1151-1164.
Service, M., 1998. Recovery of benthic communities in Strangford Lough following changes in fishing practice. ICES Council Meeting Paper, CM 1998/V.6, 13pp., Copenhagen: International Council for the Exploration of the Sea (ICES).
Shick, J.M., 2012. A functional biology of sea anemones: Springer Science & Business Media.
Shumway, S., 1978. Respiration, pumping activity and heart rate in Ciona intestinalis exposed to fluctuating salinities. Marine Biology, 48 (3), 235-242.
Shumway, S.E., 1978. Activity and respiration of the sea anemone, Metridium senile (L.) exposed to salinity fluctuations. Journal of Experimental Marine Biology and Ecology, 33, 85-92.
Smith, J.E. (ed.), 1968. 'Torrey Canyon'. Pollution and marine life. Cambridge: Cambridge University Press.
Stanley, J.A., Wilkens, S., McDonald, J.I. & Jeffs, A.G., 2016. Vessel noise promotes hull fouling. In The Effects of Noise on Aquatic Life II: Springer, pp. 1097-1104.
Steele-Petrović, H.M., 1975. An explanation for the tolerance of brachiopods and relative intolerance of filter-feeding bivalves for soft muddy bottoms. Journal of Paleontology, 552-556.
Steele-Petrović, H.M., 1976. Brachiopod food and feeding processes. Palaeontology, 19 (3), 417-436
Steele-Petrović, H.M., 1979. The physiological differences between articulate brachiopods and filter-feeding bivalves as a factor in the evolution of marine level-bottom communities. Palaeontology, 22 (1), 101-134.
Svane, I., 1984. Observations on the long-term population dynamics of the perennial ascidian, Ascidia mentula O F Müller, on the Swedish west coast. The Biological Bulletin, 167 (3), 630-646.
Svane, I. & Havenhand, J.N., 1993. Spawning and dispersal in Ciona intestinalis (L.) Marine Ecology, Pubblicazioni della Stazione Zoologica di Napoli. I, 14 , 53-66.
Thayer, C.W., 1986. Are brachiopods better than bivalves? Mechanisms of turbidity tolerance and their interaction with feeding in articulates. Paleobiology, 12 (02), 161-174.
Therriault, T.W. & Herborg, L.-M., 2008. Predicting the potential distribution of the vase tunicate Ciona intestinalis in Canadian waters: informing a risk assessment. ICES Journal of Marine Science: Journal du Conseil, 65 (5), 788-794.
Thieltges, D.W., Strasser, M. & Reise, K., 2003. The American slipper-limpet Crepidula fornicata (L.) in the Northern Wadden Sea 70 years after its introduction. Helgoland Marine Research, 57, 27-33
Tillin, H. & Tyler-Walters, H., 2014b. Assessing the sensitivity of subtidal sedimentary habitats to pressures associated with marine activities. Phase 2 Report – Literature review and sensitivity assessments for ecological groups for circalittoral and offshore Level 5 biotopes. JNCC Report No. 512B, 260 pp. Available from: www.marlin.ac.uk/publications
Tillin, H.M., Kessel, C., Sewell, J., Wood, C.A. & Bishop, J.D.D., 2020. Assessing the impact of key Marine Invasive Non-Native Species on Welsh MPA habitat features, fisheries and aquaculture. NRW Evidence Report. Report No: 454. Natural Resources Wales, Bangor, 260 pp. Available from https://naturalresourceswales.gov.uk/media/696519/assessing-the-impact-of-key-marine-invasive-non-native-species-on-welsh-mpa-habitat-features-fisheries-and-aquaculture.pdf
Tunnicliffe, V. & Wilson, K., 1988. Brachiopod populations: Distribution in fjords of British Columbia(Canada) and tolerance of low oxygen concentrations. Marine ecology progress series. 47 (2), 117-128.
Van Ofwegen, L., 2015. Sarcodictyon roseum. World Register of Marine Species: [17/03/16]. Available from http://marinespecies.org/aphia.php?p=taxdetails&id=125340
Van Ofwegen, L., Grasshoff, M. & Van der Land, J., 2001. Octocorallia (excl. Pennatulacea). European register of marine species: a check-list of the marine species in Europe and a bibliography of guides to their identification. Collection Patrimoines Naturels, 50, 104-105.
Veale, L.O., Hill, A.S., Hawkins, S.J. & Brand, A.R., 2000. Effects of long term physical disturbance by scallop fishing on subtidal epifaunal assemblages and habitats. Marine Biology, 137, 325-337.
Verhoeven, J. & Van Vierssen, W., 1978a. Structure of macrophyte dominated communities in two brackish lagoons on the island of Corsica, France. Aquatic Botany, 5, 77-86.
Whittingham, D.G., 1967. Light-induction of shedding of gametes in Ciona intestinalis and Morgula manhattensis. Biological Bulletin, Marine Biological Laboratory, Woods Hole, 132, 292-298.
Wood. C., 2005. Seasearch guide to sea anemones and corals of Britain and Ireland. Ross-on-Wye: Marine Conservation Society.
WoRMS, 2015. World Register of Marine Species. (11/04/2007). http://www.marinespecies.org
Yamaguchi, M., 1975. Growth and reproductive cycles of the marine fouling ascidians Ciona intestinalis, Styela plicata, Botrylloides violaceus, and Leptoclinum mitsukurii at Aburatsubo-Moroiso Inlet (Central Japan). Marine Biology, 29 (3), 253-259.
Zahn, R., Zahn, G., Müller, W., Kurelec, B., Rijavec, M., Batel, R. & Given, R., 1981. Assessing consequences of marine pollution by hydrocarbons using sponges as model organisms. Science of The Total Environment, 20 (2), 147-169.
Zhang, J. & Fang, J., 1999. Study on the oxygen consumption rates of some common species of ascidian. Journal of fishery sciences of China, 7 (1), 16-19.
Zhang, J., Fang, J. & Dong, S., 1999. Study on the ammonia excretion rates of four species ascidian. Marine Fisheries Research, 21 (1), 31-36.
Citation
This review can be cited as:
Last Updated: 06/12/2023
