Dahlia anemone (Urticina felina)

Urticina felina at Scapa Flow, Orkneys.
Photographer: Robert Keen Copyright: Robert Keen

Urticina felina
Photographer: Dr Keith Hiscock Copyright: Dr Keith Hiscock

Urticina felina
Photographer: Cathal McNaughton Copyright: Cathal McNaughton

Urticina felina
Photographer: Cathal McNaughton Copyright: Cathal McNaughton

Urticina felina
Photographer: Julian Madle Copyright: Julian Madle

Urticina felina (showing colour variation between the two)
Photographer: Julian Madle Copyright: Julian Madle
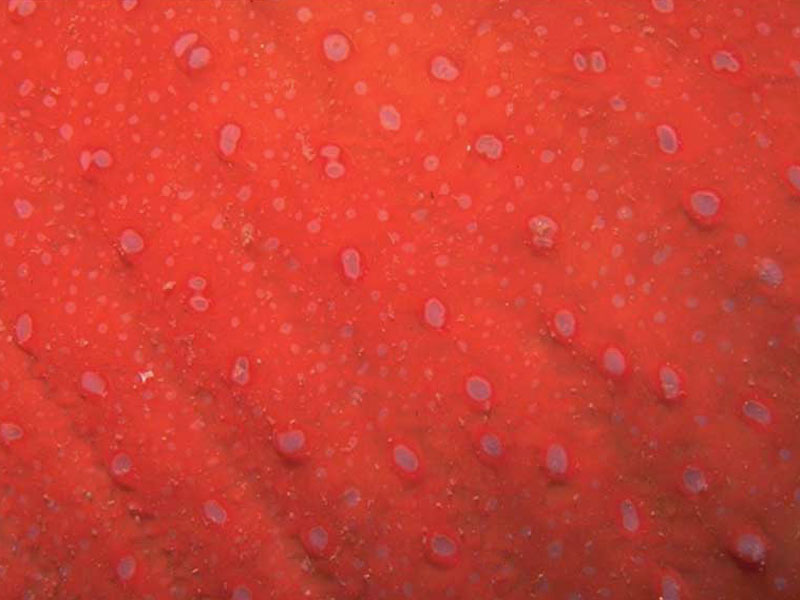
Urticina felina column showing verrucae.
Photographer: Sue Scott Copyright: Sue Scott

Urticina felina.
Photographer: Sue Scott Copyright: Sue Scott

Urticina felina.
Photographer: Sue Scott Copyright: Sue Scott

Urticina felina.
Photographer: Dr Keith Hiscock Copyright: Dr Keith Hiscock

A blue colour morph of Urticina felina.
Photographer: Paul Newland Copyright: Paul Newland

Closed Urticina felina.
Photographer: Judith Oakley Copyright: Judith Oakley

Group of Urticina felina.
Photographer: Paul Naylor Copyright: Paul Naylor

The dahlia anemone Urticina felina fully retracted. Column showing characteristic warts (verrucae) and adherent debris.
Photographer: Paul Newland Copyright: Paul Newland
Distribution data supplied by the Ocean Biodiversity Information System (OBIS). To interrogate UK data visit the NBN Atlas.Map Help
| Researched by | Angus Jackson & Dr Keith Hiscock | Refereed by | Prof. Daphne Fautin |
| Authority | (Linnaeus, 1761) | ||
| Other common names | - | Synonyms | Tealia felina (Linnaeus, 1761) |
Summary
Description
A large anemone (base up to 15 cm diameter) with up to 160 short (up to 2 cm), stout tentacles arranged in multiples of ten. Individuals from offshore tend to be larger. The coloration is very variable, ranging through white, yellow, orange, red, blue, grey, purple and brown being either plain or more commonly in some combination. Perhaps most commonly with a red column blotched with green/grey and a prominent pattern of red lines amongst the tentacle bases. The tentacles are usually banded but may be plain. There are numerous grey warts on the column to which gravel and shell fragments stick. When the tentacles are fully retracted, the body of the anemones may be almost obscured by these adherent particles.
Recorded distribution in Britain and Ireland
Found on all coasts of the British Isles.Global distribution
The species is boreal-arctic with a possible circumpolar distribution. Found throughout Europe from northern Russia to Biscay but not in the Mediterranean. Records from elsewhere are incomplete and there is considerable confusion in taxonomy.Habitat
Typically found on the lower shore and subtidally, particularly on shores with strong wave action or subtidal areas with strong tidal streams. Small individuals may be found as high as the mid-tide line. Attaches very firmly to rocks and boulders, typically in crevices and gullies, sometimes forming dense carpets. Occurs in estuaries where hard substrata are present.Depth range
down to at least 100mIdentifying features
- Large size, up to 15 cm across the base (bigger than most other anemones).
- Large verrucae or warts present on the column, often with gravel or shell fragments attached.
- Up to 160 short stout tentacles arranged in multiples of ten.
Additional information
The taxonomy and relationships of this sea anemone are in some confusion with anemones of very similar appearance and apparently reproductive biology to Urticina felina occurring on the north-west (Pacific) coast of north America. An attempt is made below to establish relationships important for using literature to support sensitivity and recoverability assessments elsewhere in this review. Stephenson (1935) identifies "Tealia (=Urticina) crassicornis" of Müller as a variety (crassicornis) of Tealia (=Urticina) felina (L.) but not the variety coriacea which is the "Tealia crassicornis" of Gosse (1860). However, Stephenson notes that, in his "var. crassicornis", the embryos develop up to a late stage in the coelenteron of the parent and later describes it as "viviparity". Since Appelöff (1900) cited in Chia & Spaulding (1972) reported that, in Europe, Tealia (=Urticina) crassicornis releases it's gametes freely into the sea (i.e. is not viviparous) and that the species they studied in the northwest USA similarly produced ova and sperm, it seems likely that their "Tealia crassicornis" has closer affinities to the British "Urticina felina" than to the species that occurs further north of the British Isles and is called "Tealia crassicornis (Müller)".
Listed by
- none -
Biology review
Taxonomy
| Level | Scientific name | Common name |
|---|---|---|
| Phylum | Cnidaria | Sea anemones, corals, sea firs & jellyfish |
| Class | Anthozoa | Sea anemones, soft & cup corals, sea pens & sea pansies |
| Order | Actiniaria | |
| Family | Actiniidae | |
| Genus | Urticina | |
| Authority | (Linnaeus, 1761) | |
| Recent Synonyms | Tealia felina (Linnaeus, 1761) | |
Biology
| Parameter | Data | ||
|---|---|---|---|
| Typical abundance | Moderate density | ||
| Male size range | up to 150mm | ||
| Male size at maturity | |||
| Female size range | Medium(11-20 cm) | ||
| Female size at maturity | |||
| Growth form | Globose | ||
| Growth rate | Data deficient | ||
| Body flexibility | High (greater than 45 degrees) | ||
| Mobility | |||
| Characteristic feeding method | Non-feeding, Predator | ||
| Diet/food source | |||
| Typically feeds on | See Additional Information. | ||
| Sociability | No information | ||
| Environmental position | Epibenthic | ||
| Dependency | No information found. | ||
| Supports | No information | ||
| Is the species harmful? | Yes The stinging nematocysts in the anemones tentacles are used to trap and paralyse prey. The nematocysts can also provoke itching and blistering of the skin in humans but the effects vary considerably between individual humans. | ||
Biology information
Densities vary from solitary individuals to dense carpets in ideal locations such as crevices and gullies. Measurements of size refer to the diameter across the base. Growth is dependent on the level of feeding so size is not proportional to age. Gosse (1860) notes [most likely from aquarium observations] that "the shore crab (Carcinus) is its ordinary prey but it feeds on limpets, and other Mollusca and nereids and shrimps and on Echinus [now Psammechinus] miliaris. Rasmussen (1973) records Urticina felina as feeding mainly on gammarids in banks of Mytilus edulis.
Habitat preferences
| Parameter | Data |
|---|---|
| Physiographic preferences | Open coast, Offshore seabed, Strait or Sound, Sea loch or Sea lough, Ria or Voe, Estuary |
| Biological zone preferences | Lower circalittoral, Lower eulittoral, Lower infralittoral, Sublittoral fringe, Upper circalittoral, Upper infralittoral |
| Substratum / habitat preferences | Bedrock, Crevices / fissures, Large to very large boulders, Other species, Small boulders |
| Tidal strength preferences | Moderately strong 1 to 3 knots (0.5-1.5 m/sec.), Strong 3 to 6 knots (1.5-3 m/sec.), Very strong > 6 knots (>3 m/sec.), Weak < 1 knot (<0.5 m/sec.) |
| Wave exposure preferences | Exposed, Extremely exposed, Moderately exposed, Sheltered, Very exposed |
| Salinity preferences | Full (30-40 psu), Low (<18 psu), Reduced (18-30 psu), Variable (18-40 psu) |
| Depth range | down to at least 100m |
| Other preferences | No text entered |
| Migration Pattern | Non-migratory or resident |
Habitat Information
- Individuals found further offshore tend to be larger in size.
- Urticina eques is a similar but larger species (up to 30 cm tentacle spread) with longer tentacles and more commonly found offshore and in deeper water to 400 m. This species has fewer or no verrucae and no attached gravel or other particles.
- Rasmussen, (1973) records Urticina felina as being very common in banks of Mytilus feeding mostly on gammarids.
- Urticina felina is recorded from several estuarine sites including Mucking in Thames estuary and the river Blackwater estuary and so will be subject to variable or low salinities. In the Westerschelde estuary, Braber & Borghouts (1977) found that Urticina (as Tealia) felina penetrated to about the 11ppt Chlorinity (about 20 psu) isohaline at mid tide during average water discharge making it tolerant of reduced salinity conditions.
Life history
Adult characteristics
| Parameter | Data |
|---|---|
| Reproductive type | Gonochoristic (dioecious) |
| Reproductive frequency | Annual protracted |
| Fecundity (number of eggs) | No information |
| Generation time | Insufficient information |
| Age at maturity | See additional information |
| Season | April - June |
| Life span | See additional information |
Larval characteristics
| Parameter | Data |
|---|---|
| Larval/propagule type | - |
| Larval/juvenile development | Lecithotrophic |
| Duration of larval stage | See additional information |
| Larval dispersal potential | See additional information |
| Larval settlement period | Insufficient information |
Life history information
- No information has been found regarding the longevity of Urticina felina but given the large size, slow growth rate and few predators it is likely that it survives for quite a long time. Specimens in aquarium tanks are known to still be flourishing fifty years after collection (P. G. Moore pers. comm.).
- Age at maturity is not known. Chia & Spaulding (1972) working with the similar (see 'Taxonomy') Tealia crassicornis (see below) found no sign of gonad development at 14 months old. The smallest fertile Urticina lofotensis, a similar species in California, are recorded as at least 18 months old (Wedi & Dunn, 1983).
- Solé-Cava et al. (1985) considered that sexual reproduction is the most important, if not the only, method of reproduction in Urticina felina. Appeloff (1900) (reported in Chia & Spaulding, 1972) observed that in Europe "Tealia (=Urticina) crassicornis" releases its gametes into the sea and that larval development is independent of the adult. Chia & Spaulding (1972), in observing that Tealia crassicornis from the north-west of the USA (the Pacific coast) has a mode of development similar to that described by Appeloff (almost certainly for what is now called "Urticina felina") suggests that the information they collected on Tealia crassicornis can be used with some validity here. It is not known whether Urticina felina reproduces asexually as do several other anemones (such as Actinia equina and Metridium senile). Stephenson (1935) reports that viviparity has been suspected because of the sudden appearance apparently from "nowhere" of individuals in aquaria.
- The Plymouth Marine Fauna (Marine Biological Association, 1957) records Urticina felina as breeding in May. Chia & Spaulding (1972) record the similar Tealia crassicornis from San Juan Island on the north-west coast of the USA as spawning in the morning during April, May and June.
- Chia & Spaulding (1972) bred and grew Tealia crassicornis from the north-west coast of the USA. In Tealia crassicornis, mucus containing gametes were expelled from the mouth. The yellow eggs (500-700 µm diameter) formed little clusters which then broke apart and began to float.
- The duration of the larval stage may vary. For Tealia crassicornis, Chia & Spaulding (1972) found that nine days after fertilization, the planula was ready to settle and, a further four days after settling, had 4 tentacles. Certain substrata (such as Phyllochaetopterus sp. and Sabellaria cementaria tubes) could induce settlement rapidly in the laboratory. In the absence of inducing substrata larvae could remain in the water column for at least 17 days but settled within the second month after fertilization.
- The species is probably quite slow growing. Chia & Spaulding (1972) found that fed individuals of the similar Tealia crassicornis were only 10mm in diameter after a year and there was no gonad development present in 14 month old anemones. However, at 18 months, individuals were 4 cm diameter with 60-70 tentacles.
- Solé-Cava et al. (1994) suggested that the large sub-littoral sea anemone Urticina eques (very similar to Urticina felina) with its large lecithotrophic larvae is probably not truly planktonic and has poor dispersive powers.
Sensitivity review
The MarLIN sensitivity assessment approach used below has been superseded by the MarESA (Marine Evidence-based Sensitivity Assessment) approach (see menu). The MarLIN approach was used for assessments from 1999-2010. The MarESA approach reflects the recent conservation imperatives and terminology and is used for sensitivity assessments from 2014 onwards.
Physical pressures
Use / to open/close text displayed
| Intolerance | Recoverability | Sensitivity | Evidence / Confidence | |
Substratum loss [Show more]Substratum lossBenchmark. All of the substratum occupied by the species or biotope under consideration is removed. A single event is assumed for sensitivity assessment. Once the activity or event has stopped (or between regular events) suitable substratum remains or is deposited. Species or community recovery assumes that the substratum within the habitat preferences of the original species or community is present. Further details EvidenceUrticina felina anemones adhere strongly to the substratum. Substratum loss would result in mortality. Some individuals might, however, be left behind as they typically live in fissures. Recruitment to replace lost individuals is likely to be slow (see 'Additional Information' below). | High | Moderate | Moderate | Moderate |
Smothering [Show more]SmotheringBenchmark. All of the population of a species or an area of a biotope is smothered by sediment to a depth of 5 cm above the substratum for one month. Impermeable materials, such as concrete, oil, or tar, are likely to have a greater effect. Further details. EvidenceUrticina felina anemones adhere strongly to the substratum and would be entirely covered by smothering material. However, Urticina felina lives in situations where it may be covered from time-to-time by sediment, especially coarser substrata which suggests some ability to survive. For example, Holme & Wilson (1985) observed Urticina felina attached to pebbles, cobbles or rock subject to sand scour or periodic smothering by sand at 50-55m depth, offshore, in the western English Channel. The tidal streams in the central parts of the Channel may reach 125 cm/s during neaps and 166 cm/s on springs. Therefore, he suggested that Urticina felina was tolerant of sand scour or periodic smothering by < ca. 5cm of sand, being able to extend its column to maintain its disc above the sand surface (Holme & Wilson, 1985). Thus the species is considered to have low intolerance to smothering. As the species is able to maintain its disc above the smothering material recovery is very rapid. Adults can also detach from the substratum and relocate but locomotive ability is very limited. | Low | Very high | Very Low | High |
Increase in suspended sediment [Show more]Increase in suspended sedimentBenchmark. An arbitrary short-term, acute change in background suspended sediment concentration e.g., a change of 100 mg/l for one month. The resultant light attenuation effects are addressed under turbidity, and the effects of rapid settling out of suspended sediment are addressed under smothering. Further details EvidenceBeing an epibenthic species, Urticina felina would be exposed to changes in siltation. Increases in siltation may begin to cover the anemone or interfere with feeding. An energetic cost will result from efforts to clean off the silt particles, e.g. through mucus production and sloughing. Repeated energetic expenditure in cleaning off silt particles may cause loss of condition. Recovery of condition may take several months. | Low | Very high | Very Low | Moderate |
Decrease in suspended sediment [Show more]Decrease in suspended sedimentBenchmark. An arbitrary short-term, acute change in background suspended sediment concentration e.g., a change of 100 mg/l for one month. The resultant light attenuation effects are addressed under turbidity, and the effects of rapid settling out of suspended sediment are addressed under smothering. Further details EvidenceReduction of the need to keep the anemone surface clear of silt will mean less energy expenditure and mucus production and therefore likely benefit to the anemone. | Tolerant* | Not relevant | Not sensitive* | High |
Desiccation [Show more]Desiccation
EvidenceThe species is found on the lower shore as well as subtidally. Small individuals have been recorded as high as the mid tide line (Manuel, 1988). Decreases in desiccation will have no effect. The anemone is able to detach from the substratum and relocate in order to find better or avoid unfavourable conditions. Increases in desiccation may cause part of an intertidal population to die before suitable relocation can occur. Recruitment to replace lost individuals is likely to be slow (see 'Additional Information' below). A precautionary assessment of 'moderate' recoverability is made but with a low confidence. | Intermediate | Moderate | Moderate | Low |
Increase in emergence regime [Show more]Increase in emergence regimeBenchmark. A one hour change in the time covered or not covered by the sea for a period of one year. Further details EvidenceIncrease in emergence is likely to result in exposure to desiccation and a decreased opportunity for feeding. Increased emergence may have no effect for anemones in damp fissures or pools. Assuming that desiccation or heat stress (see 'Increase in temperature' below) occurs, intolerance and recoverability will be as desiccation. Recruitment to replace lost individuals is likely to be slow (see 'Additional Information' below). A precautionary assessment of 'moderate' recoverability is made but with a low confidence. | Intermediate | Moderate | Moderate | Low |
Decrease in emergence regime [Show more]Decrease in emergence regimeBenchmark. A one hour change in the time covered or not covered by the sea for a period of one year. Further details EvidenceUrticina felina is predominantly a subtidal species so that decrease in emergence is likely to lead to more habitats for colonization becoming available. | Tolerant* | Not relevant | Not sensitive* | High |
Increase in water flow rate [Show more]Increase in water flow rateA change of two categories in water flow rate (view glossary) for 1 year, for example, from moderately strong (1-3 knots) to very weak (negligible). Further details EvidenceThe species favours areas with strong tidal currents (Holme & Wilson, 1985; Migné & Davoult, 1997) although it is also found in more calm and sheltered areas as well as deep water. The anemone is very firmly attached and, although there may be some inhibition of feeding in very strong flows, increases in water flow rate are not likely to have a significant effect on Urticina felina. | Tolerant | Not relevant | Not sensitive | Moderate |
Decrease in water flow rate [Show more]Decrease in water flow rateA change of two categories in water flow rate (view glossary) for 1 year, for example, from moderately strong (1-3 knots) to very weak (negligible). Further details EvidenceIn the absence of wave action, water flow is likely to be very important in preventing siltation and stagnation and in bringing food. Therefore, in conditions where water flow rates fall to very low levels, anemones may be adversely affected, lose condition and, especially if some stagnation occurs, some may die. Although recolonization is likely to occur from nearby populations, frequency and success of recruitment is unclear and a precautionary assessment of 'Moderate' is made (see 'Additional Information' below). | Intermediate | Moderate | Moderate | Moderate |
Increase in temperature [Show more]Increase in temperature
For intertidal species or communities, the range of temperatures includes the air temperature regime for that species or community. Further details EvidenceThe species distribution extends to the north and south of the British Isles (Manuel, 1988) and so the species is unlikely to be affected by an increase in open water temperatures. Gosse (1860) observed that Urticina felina (as Actinia crassicornis) was "one of the most difficult [anemones] to keep in an aquarium" and that "the heat of the summer is generally fatal to our captive specimens". It is therefore likely that local warming may adversely affect individuals and that some mortality might occur. Although recolonization is likely to occur from nearby populations, frequency and success of recruitment is unclear and a precautionary assessment of 'Moderate' is made (see 'Additional Information' below). | Intermediate | High | Low | Low |
Decrease in temperature [Show more]Decrease in temperature
For intertidal species or communities, the range of temperatures includes the air temperature regime for that species or community. Further details EvidenceThe species distribution extends to the north and south of the British Isles (Manuel, 1988) and so the species is unlikely to be affected by a decrease in open water temperatures. Although Urticina felina was apparently unaffected by the extremely cold winter of 1962/3 (Crisp, 1964), Gosse (1860) observed that "after the intense and protracted frost of February 1855, the shores of South Devon were strewn with dead and dying anemones, principally of this species". Bearing in mind the equivocal observations from two cold winters, it is suggested that at least some individuals might be killed by extreme cold. Although recolonization is likely to occur from nearby populations, frequency and success of recruitment is unclear and a precautionary assessment of 'Moderate' is made (see 'Additional Information' below). | Intermediate | High | Low | Low |
Increase in turbidity [Show more]Increase in turbidity
EvidenceUrticina felina occurs in clear to highly turbid waters and occurs down to depths of at least 100m (Manuel, 1988) where light levels are low. The anemone is not known to contain symbiotic algae and is likely to tolerate changes in turbidity or the resulting change in light attenuation. | Tolerant | Not relevant | Not sensitive | Low |
Decrease in turbidity [Show more]Decrease in turbidity
EvidenceUrticina felina occurs in clear to highly turbid waters and occurs down to depths of at least 100m (Manuel, 1988) where light levels are low. The anemone is not known to contain symbiotic algae and is unlikely to be sensitive to changes in turbidity or the resulting change in light attenuation. | Tolerant | Not relevant | Not sensitive | High |
Increase in wave exposure [Show more]Increase in wave exposureA change of two ranks on the wave exposure scale (view glossary) e.g., from Exposed to Extremely exposed for a period of one year. Further details EvidenceThe species favours areas with strong wave action (Manuel, 1988) and strong tidal currents (Migné & Davoult, 1997) although it is also found in more calm and sheltered areas as well as deep water. | Tolerant | Not relevant | Not sensitive | Low |
Decrease in wave exposure [Show more]Decrease in wave exposureA change of two ranks on the wave exposure scale (view glossary) e.g., from Exposed to Extremely exposed for a period of one year. Further details EvidenceIn the absence of tidal streams, wave action is likely to be very important in preventing siltation and stagnation and in bringing food. Therefore, in conditions where wave action falls to very low levels, anemones may be adversely affected, lose condition and, especially if some stagnation occurs, some may die. Assuming that some individuals survive, local recruitment is likely to occur within a few years. | Intermediate | High | Low | Moderate |
Noise [Show more]Noise
EvidenceUrticina felina is likely to have poor ability for detection of noise vibrations and as such is unlikely to be sensitive to noise. | Tolerant | Not relevant | Not sensitive | Low |
Visual presence [Show more]Visual presenceBenchmark. The continuous presence for one month of moving objects not naturally found in the marine environment (e.g., boats, machinery, and humans) within the visual envelope of the species or community under consideration. Further details EvidenceUrticina felina has very limited, if any, ability for visual perception. The anemone is unlikely to be sensitive to visual presence. | Tolerant | Not relevant | Not sensitive | High |
Abrasion & physical disturbance [Show more]Abrasion & physical disturbanceBenchmark. Force equivalent to a standard scallop dredge landing on or being dragged across the organism. A single event is assumed for assessment. This factor includes mechanical interference, crushing, physical blows against, or rubbing and erosion of the organism or habitat of interest. Where trampling is relevant, the evidence and trampling intensity will be reported in the rationale. Further details. EvidenceThe species occurs frequently in areas with strong tidal currents and coarse sediments and so may be exposed to and tolerant of particle scour. The anemone is also soft, flexible and can reform its attachment to the substratum. The anemone lives in fissures, which may be protected from abrasive forces. However, physical impact is likely to cause damage and mortality to exposed individuals. It is therefore suggested that some individuals may be killed by a physical disturbance event. Although recolonization is likely to occur from nearby populations, frequency, and success of recruitment is unclear and a precautionary assessment of 'Moderate' is made (see additional information below). | Intermediate | Moderate | Moderate | Low |
Displacement [Show more]DisplacementBenchmark. Removal of the organism from the substratum and displacement from its original position onto a suitable substratum. A single event is assumed for assessment. Further details EvidenceAlthough Urticina felina adheres strongly to the substratum, it does not form a permanent attachment. The anemones can move around to re-attach in a better position. Displacement may result in loss of a good position or hinder feeding but will only have serious consequences if the basal tissue is damaged - as often happens when anemones are collected for the aquarium. The assessment made here assumes no damage to the tissue. Relocation to a suitable area will be accompanied by recovery. | Low | Immediate | Not sensitive | Low |
Chemical pressures
Use [show more] / [show less] to open/close text displayed
| Intolerance | Recoverability | Sensitivity | Evidence / Confidence | |
Synthetic compound contamination [Show more]Synthetic compound contaminationSensitivity is assessed against the available evidence for the effects of contaminants on the species (or closely related species at low confidence) or community of interest. For example:
The evidence used is stated in the rationale. Where the assessment can be based on a known activity then this is stated. The tolerance to contaminants of species of interest will be included in the rationale when available; together with relevant supporting material. Further details. EvidenceVery little information has been found. Hoare & Hiscock (1974) observed that Urticina felina survived near to an acidified halogenated effluent discharge in a 'transition' zone where many other species were unable to survive, suggesting a tolerance to chemical contamination. However, Urticina felina was absent from stations closest to the effluent which were dominated by pollution tolerant species particularly polychaetes. Those specimens closest to the effluent discharge appeared generally unhealthy. Because it appears that Urticina was unable to tolerate the most polluted conditions, intolerance has been assessed as intermediate. Although recolonization is likely to occur from nearby populations, frequency and success of recruitment is unclear and a precautionary assessment of 'Moderate' is made (see 'Additional Information' below). | Intermediate | Moderate | Moderate | Low |
Heavy metal contamination [Show more]Heavy metal contaminationEvidenceInsufficientinformation. | No information | No information | No information | Not relevant |
Hydrocarbon contamination [Show more]Hydrocarbon contaminationEvidenceOne month after the Torrey Canyon oil spill the dahlia anemone, Urticina felina, was found to be one of the most resistant animals on the shore, being commonly found alive in pools between the tide-marks which appeared to be devoid of all other animals (Smith, 1968). Intolerance is, therefore, assessed as low for dispersed or liquid oil. 'Condition' would be likely to return to normal once the oil is removed. However, the species may be susceptible to smothering effects and, in the case of thick oil, mortality seems likely. | Low | Very high | Very Low | Moderate |
Radionuclide contamination [Show more]Radionuclide contaminationEvidenceInsufficientinformation | No information | No information | No information | Not relevant |
Changes in nutrient levels [Show more]Changes in nutrient levelsEvidenceInsufficientinformation | No information | No information | No information | Not relevant |
Increase in salinity [Show more]Increase in salinity
EvidenceUrticina felina lives in full salinity situations and the factor is assessed as Not Relevant. | Not relevant | Not relevant | Not relevant | Not relevant |
Decrease in salinity [Show more]Decrease in salinity
EvidenceAlthough Urticina felina is predominantly marine, the species does penetrate into estuaries (e.g. the Thames estuary at Mucking (NMMP, 2001) and the River Blackwater estuary (Davis, 1967). Braber & Borghouts (1977) found that Urticina (as Tealia) felina penetrated to about the 11ppt Chlorinity (about 20psu) isohaline at mid tide during average water discharge in the Westerschelde estuary suggesting that, during high river flow, it would be tolerant of reduced salinity conditions. Intertidal and rock pool individuals will also be subject to variations in salinity because of precipitation on the shore; albeit for short periods on the lower shore. Therefore, the species seems to have a high tolerance to reduction in salinity but may have to retract tentacles and suffer reduced opportunity to feed. Intolerance has therefore been assessed as low suggesting that individuals are unlikely to be killed by changes at the level of the benchmark. Recovery is in terms of condition and is therefor very high. | Low | Very high | Very Low | Moderate |
Changes in oxygenation [Show more]Changes in oxygenationBenchmark. Exposure to a dissolved oxygen concentration of 2 mg/l for one week. Further details. EvidenceThere is no information about Urticina felina tolerance to changes in oxygenation but Cole et al., (1999) suggest possible adverse effects on marine species below 4 mg/l and probable adverse effects below 2mg/l. The large size and slow growth rate of this anemone suggests that it is quite long lived. Although recolonization is likely to occur from nearby populations, frequency and success of recruitment is unclear and a precautionary assessment of 'Moderate' is made (see 'Additional Information' below). | Intermediate | High | Low | Low |
Biological pressures
Use [show more] / [show less] to open/close text displayed
| Intolerance | Recoverability | Sensitivity | Evidence / Confidence | |
Introduction of microbial pathogens/parasites [Show more]Introduction of microbial pathogens/parasitesBenchmark. Sensitivity can only be assessed relative to a known, named disease, likely to cause partial loss of a species population or community. Further details. EvidenceInsufficientinformation | No information | No information | No information | Not relevant |
Introduction of non-native species [Show more]Introduction of non-native speciesSensitivity assessed against the likely effect of the introduction of alien or non-native species in Britain or Ireland. Further details. EvidenceNo known non-native species compete with Urticina felina. | No information | No information | No information | Not relevant |
Extraction of this species [Show more]Extraction of this speciesBenchmark. Extraction removes 50% of the species or community from the area under consideration. Sensitivity will be assessed as 'intermediate'. The habitat remains intact or recovers rapidly. Any effects of the extraction process on the habitat itself are addressed under other factors, e.g. displacement, abrasion and physical disturbance, and substratum loss. Further details. EvidenceUrticina felina is not currently subject to extraction. However if a cold water marine aquarium trade were to take-off, this species is likely to be collected. Although Urticina felina probably breeds every year there is no information regarding fecundity. Although recolonization is likely to occur from nearby populations, frequency and success of recruitment is unclear and a precautionary assessment of 'Moderate' is made. See 'Additional Information' below. | High | Moderate | Moderate | Moderate |
Extraction of other species [Show more]Extraction of other speciesBenchmark. A species that is a required host or prey for the species under consideration (and assuming that no alternative host exists) or a keystone species in a biotope is removed. Any effects of the extraction process on the habitat itself are addressed under other factors, e.g. displacement, abrasion and physical disturbance, and substratum loss. Further details. EvidenceUrticina felina has no known obligate relationships so removal of other species is unlikely to have any direct effect. The incidental physical effects of removal of other species can be assessed under the relevant factors. | Tolerant | Not relevant | Not sensitive | Low |
Additional information
Recoverability is likely to be slow in populations where nearby individuals do not exist. The large size, slow growth rate and evidence from aquarium populations suggests that Urticina felina is long lived. Although it probably breeds each year there is no information regarding fecundity. Breeding probably does not occur until the anemone is at least 1.5 years old. Dispersal ability is considered to be poor in the similar Urticina eques (Solé-Cava et al., 1994). The larva is most likely benthic and, although unlikely to settle for many days after release (based on work on the similar Tealia crassicornis for north-west USA), is unlikely to travel far. However, assuming that there are populations surviving nearby (further down the shore), recruitment is likely to occur over the short distances involved but how rapidly is uncertain. Adults can detach from the substratum and relocate but locomotive ability is very limited. There is potential for some immigration of adults from other populations via water currents or rafting. Gosse (1853) noted that an Eolis papillosa had "eaten a hole the size of a pea in the side before being discovered". Under its current name of %Aeolidia papillosa%, Reidy (1996) describes how the plumose anemone %Metridium senile% is preferred as a food source to %Urticina crassicornis% but that newly settled Aeolidea papillosa occurred especially adjacent to individuals of Urticina crassicornis. (Urticina crassicornis is a similar species to Urticina felina)Importance review
Policy/legislation
- no data -
Status
| National (GB) importance | - | Global red list (IUCN) category | - |
Non-native
| Parameter | Data |
|---|---|
| Native | - |
| Origin | - |
| Date Arrived | - |
Importance information
In ideal conditions such as crevices and gullies on wave exposed shores the species can form dense carpets. The role of Urticina sea anemones as refuges and feeding stations for the painted greenling Oxylebius pictus (NE Pacific) has been investigated by Elliott (1992). The same roles may be filled by species in the British Isles although there are no records of this for Urticina felina. In some strong tidal current communities, Urticina felina, along with Alcyonium digitatum and Ophiothrix fragilis can constitute a large proportion of the biomass. In addition, they are thought to be responsible for the major part of carbon / nitrogen exchange at the sediment - water boundary (Migné & Davoult, 1995; Migné & Davoult, 1997a). Although 'Culinary use' is indicated as 'no', Gosse (1853) describes methods of cooking the dahlia anemone and seemed reasonably impressed with it boiled or fried.Bibliography
Braber, L. & Borghouts, C.H., 1977. Distribution and ecology of Anthozoa in the estuarine region of the rivers Rhine, Meuse and Scheldt. Hydrobiologia, 52, 15-21.
Bruce, J.R., Colman, J.S. & Jones, N.S., 1963. Marine fauna of the Isle of Man. Liverpool: Liverpool University Press.
Chia, F.S. & Spaulding, J.G., 1972. Development and juvenile growth of the sea anemone Tealia crassicornis. Biological Bulletin, Marine Biological Laboratory, Woods Hole, 142, 206-218.
Cole, S., Codling, I.D., Parr, W. & Zabel, T., 1999. Guidelines for managing water quality impacts within UK European Marine sites. Natura 2000 report prepared for the UK Marine SACs Project. 441 pp., Swindon: Water Research Council on behalf of EN, SNH, CCW, JNCC, SAMS and EHS. [UK Marine SACs Project.]. Available from: http://ukmpa.marinebiodiversity.org/uk_sacs/pdfs/water_quality.pdf
Crisp, D.J. (ed.), 1964. The effects of the severe winter of 1962-63 on marine life in Britain. Journal of Animal Ecology, 33, 165-210.
Davis, D.S., 1967. The marine fauna of the Blackwater Estuary and adjacent waters. Essex Naturalist, 32, 1-60.
Elliott, J., 1992. The role of sea anemones as refuges and feeding habitats for the temperate fish Oxylebius pictus. Environmental Biology of Fishes, 35, 381-400.
Eno, N.C., Clark, R.A. & Sanderson, W.G. (ed.) 1997. Non-native marine species in British waters: a review and directory. Peterborough: Joint Nature Conservation Committee.
George, J.D., Tittley, I., Price, J.H., & Fincham, A.A., 1988. The macrobenthos of chalk shores in North Norfolk and around Flamborough Headland (North Humberside). Nature Conservancy Council CSD Rep. 833 149p., Peterborough: Nature Conservancy Council
Gosse, P.H., 1853. A naturalist's rambles on the Devonshire coast. London: Van Voorst.
Gosse, P.H., 1860. Actinologia Britannica: a history of British sea anemones and corals. London: Van Voorst.
Hand, C., 1955b. The sea anemones of central California. Part II. The endomyarian and mesomyarian anemones. The Wassmann Journal of Biology, 13, 37-99.
Hayward, P., Nelson-Smith, T. & Shields, C. 1996. Collins pocket guide. Sea shore of Britain and northern Europe. London: HarperCollins.
Hoare, R. & Hiscock, K., 1974. An ecological survey of the rocky coast adjacent to the effluent of a bromine extraction plant. Estuarine and Coastal Marine Science, 2 (4), 329-348.
Holme, N.A. & Wilson, J.B., 1985. Faunas associated with longitudinal furrows and sand ribbons in a tide-swept area in the English Channel. Journal of the Marine Biological Association of the United Kingdom, 65, 1051-1072.
Houtman, R., Paul, L.R., Ungemach, R.V., & Ydenberg, R.C., 1997. Feeding and predator avoidance in the rose anemone Urticina piscivora. Marine Biology, 128, 225-229.
Howson, C.M. & Picton, B.E., 1997. The species directory of the marine fauna and flora of the British Isles and surrounding seas. Belfast: Ulster Museum. [Ulster Museum publication, no. 276.]
JNCC (Joint Nature Conservation Committee), 1999. Marine Environment Resource Mapping And Information Database (MERMAID): Marine Nature Conservation Review Survey Database. [on-line] http://www.jncc.gov.uk/mermaid
Manuel, R.L., 1988. British Anthozoa. Synopses of the British Fauna (New Series) (ed. D.M. Kermack & R.S.K. Barnes). The Linnean Society of London [Synopses of the British Fauna No. 18.]. DOI https://doi.org/10.1002/iroh.19810660505
MBA (Marine Biological Association), 1957. Plymouth Marine Fauna. Plymouth: Marine Biological Association of the United Kingdom.
Meyling, A.W. Gmelig, 1998. Zeedahlia - Urticina felina (Linnaeus, 1761) http://wiebrand.www.cistron.nl/anemonen/urtfel.htm, 2000-09-01
Migné, A., & Davoult, D., 1995. Role des organismes suspensivores dans les transferts pélago-benthiques d'une zone de fort hydrodynamisme: approche expérimentale. Journal de Recherche Oceanographique, 20, 9-14.
Migné, A., & Davoult, D., 1997a. Ammonium excretion in two benthic cnidarians: Alcyonium digitatum (Linnaeus, 1758) and Urticina felina (Linnaeus, 1767). Journal of Sea Research, 37, 101-107.
Moen, F.E., 1996. Urticina felina Fjaeresjorose.
NMMP, 2001. National Marine Monitoring Programme (NMMP). http://www.marlin.ac.uk, 2001-01-17
Picton, B.E. & Costello, M.J., 1998. BioMar biotope viewer: a guide to marine habitats, fauna and flora of Britain and Ireland. [CD-ROM] Environmental Sciences Unit, Trinity College, Dublin.
Rasmussen, E., 1973. Systematics and ecology of the Isefjord marine fauna (Denmark). Ophelia, 11, 1-507.
Reidy, S. 1996. Comparison of associations of the nudibranch Aeolidia papillosa with two sea anemones Urticina crassicornis and Metridium senile. In Proceedings of the 24th Annual Benthic Ecology Meeting, Columbia, South Carolina, March 7-10, 1996 (ed. S.A. Woodin et al.), pp. 68.
Smith, J.E. (ed.), 1968. 'Torrey Canyon'. Pollution and marine life. Cambridge: Cambridge University Press.
Solé-Cava, A.M. & Thorpe, J.P., 1992. Genetic divergence between colour morphs in populations of the common intertidal sea anemones Actinia equina and A. prasina (Anthozoa: Actiniaria). Marine Biology, 112, 243-252.
Solé-Cava, A.M., Thorpe, J.P. & Kay, J.G., 1985. Reproductive isolation with little genetic divergence between Urticina (= Tealia) felina and U. eques (Anthozoa: Actiniaria). Marine Biology, 85, 279-284.
Solé-Cava, A.M., Thorpe, J.P. & Todd, C.D., 1994. High genetic similarity between geographically distant populations in a sea anemone with low dispersal capabilities. Journal of the Marine Biological Association of the United Kingdom, 74, 895-902.
Spaulding, J.G., 1974. Embryonic and larval development in sea anemones (Anthozoa: Actiniaria). American Zoologist, 14, 511-520.
Stephenson, T.A., 1935. The British Sea Anemones, vol. 2. London: Ray Society.
Wedi, S.E., & Dunn, D.F., 1983. Gametogenesis and reproductive periodicity of the subtidal sea anemone Urticina lofotensis (Coelenterata: Actiniaria) in California. Biological Bulletin, Marine Biological Laboratory, Woods Hole, 165, 458-472.
Datasets
Bristol Regional Environmental Records Centre, 2017. BRERC species records recorded over 15 years ago. Occurrence dataset: https://doi.org/10.15468/h1ln5p accessed via GBIF.org on 2018-09-25.
Bristol Regional Environmental Records Centre, 2017. BRERC species records within last 15 years. Occurrence dataset: https://doi.org/10.15468/vntgox accessed via GBIF.org on 2018-09-25.
Centre for Environmental Data and Recording, 2018. IBIS Project Data. Occurrence dataset: https://www.nmni.com/CEDaR/CEDaR-Centre-for-Environmental-Data-and-Recording.aspx accessed via NBNAtlas.org on 2018-09-25.
Centre for Environmental Data and Recording, 2018. Ulster Museum Marine Surveys of Northern Ireland Coastal Waters. Occurrence dataset https://www.nmni.com/CEDaR/CEDaR-Centre-for-Environmental-Data-and-Recording.aspx accessed via NBNAtlas.org on 2018-09-25.
Cofnod – North Wales Environmental Information Service, 2018. Miscellaneous records held on the Cofnod database. Occurrence dataset: https://doi.org/10.15468/hcgqsi accessed via GBIF.org on 2018-09-25.
Environmental Records Information Centre North East, 2018. ERIC NE Combined dataset to 2017. Occurrence dataset: http://www.ericnortheast.org.ukl accessed via NBNAtlas.org on 2018-09-38
Fenwick, 2018. Aphotomarine. Occurrence dataset http://www.aphotomarine.com/index.html Accessed via NBNAtlas.org on 2018-10-01
Fife Nature Records Centre, 2018. St Andrews BioBlitz 2014. Occurrence dataset: https://doi.org/10.15468/erweal accessed via GBIF.org on 2018-09-27.
Fife Nature Records Centre, 2018. St Andrews BioBlitz 2015. Occurrence dataset: https://doi.org/10.15468/xtrbvy accessed via GBIF.org on 2018-09-27.
Fife Nature Records Centre, 2018. St Andrews BioBlitz 2016. Occurrence dataset: https://doi.org/10.15468/146yiz accessed via GBIF.org on 2018-09-27.
Isle of Wight Local Records Centre, 2017. IOW Natural History & Archaeological Society Marine Invertebrate Records 1853- 2011. Occurrence dataset: https://doi.org/10.15468/d9amhg accessed via GBIF.org on 2018-09-27.
Kent Wildlife Trust, 2018. Kent Wildlife Trust Shoresearch Intertidal Survey 2004 onwards. Occurrence dataset: https://www.kentwildlifetrust.org.uk/ accessed via NBNAtlas.org on 2018-10-01.
Manx Biological Recording Partnership, 2017. Isle of Man wildlife records from 01/01/2000 to 13/02/2017. Occurrence dataset: https://doi.org/10.15468/mopwow accessed via GBIF.org on 2018-10-01.
Manx Biological Recording Partnership, 2018. Isle of Man historical wildlife records 1990 to 1994. Occurrence dataset: https://doi.org/10.15468/aru16v accessed via GBIF.org on 2018-10-01.
Manx Biological Recording Partnership, 2018. Isle of Man historical wildlife records 1995 to 1999. Occurrence dataset: https://doi.org/10.15468/lo2tge accessed via GBIF.org on 2018-10-01.
Manx Biological Recording Partnership, 2022. Isle of Man historical wildlife records 1990 to 1994. Occurrence dataset:https://doi.org/10.15468/aru16v accessed via GBIF.org on 2024-09-27.
National Trust, 2017. National Trust Species Records. Occurrence dataset: https://doi.org/10.15468/opc6g1 accessed via GBIF.org on 2018-10-01.
NBN (National Biodiversity Network) Atlas. Available from: https://www.nbnatlas.org.
North East Scotland Biological Records Centre, 2017. NE Scotland other invertebrate records 1800-2010. Occurrence dataset: https://doi.org/10.15468/ifjfxz accessed via GBIF.org on 2018-10-01.
OBIS (Ocean Biodiversity Information System), 2025. Global map of species distribution using gridded data. Available from: Ocean Biogeographic Information System. www.iobis.org. Accessed: 2025-07-19
Outer Hebrides Biological Recording, 2018. Invertebrates (except insects), Outer Hebrides. Occurrence dataset: https://doi.org/10.15468/hpavud accessed via GBIF.org on 2018-10-01.
South East Wales Biodiversity Records Centre, 2018. Dr Mary Gillham Archive Project. Occurance dataset: http://www.sewbrec.org.uk/ accessed via NBNAtlas.org on 2018-10-02
South East Wales Biodiversity Records Centre, 2023. SEWBReC Marine and other Aquatic Invertebrates (South East Wales). Occurrence dataset:https://doi.org/10.15468/zxy1n6 accessed via GBIF.org on 2024-09-27.
The Wildlife Information Centre, 2018. TWIC Biodiversity Field Trip Data (1995-present). Occurrence dataset: https://doi.org/10.15468/ljc0ke accessed via GBIF.org on 2018-10-02.
Yorkshire Wildlife Trust, 2018. Yorkshire Wildlife Trust Shoresearch. Occurrence dataset: https://doi.org/10.15468/1nw3ch accessed via GBIF.org on 2018-10-02.
Citation
This review can be cited as:
Last Updated: 29/04/2008
